
Abstract
Subseafloor microbes beneath active hydrothermal vents are thought to live near the upper temperature limit for life on Earth. We drilled and cored the Iheya North hydrothermal field in the Mid-Okinawa Trough, and examined the phylogenetic compositions and the products of metabolic functions of sub-vent microbial communities. We detected microbial cells, metabolic activities and molecular signatures only in the shallow sediments down to 15.8 m below the seafloor at a moderately distant drilling site from the active hydrothermal vents (450 m). At the drilling site, the profiles of methane and sulfate concentrations and the δ13C and δD isotopic compositions of methane suggested the laterally flowing hydrothermal fluids and the in situ microbial anaerobic methane oxidation. In situ measurements during the drilling constrain the current bottom temperature of the microbially habitable zone to ~45 °C. However, in the past, higher temperatures of 106–198 °C were possible at the depth, as estimated from geochemical thermometry on hydrothermally altered clay minerals. The 16S rRNA gene phylotypes found in the deepest habitable zone are related to those of thermophiles, although sequences typical of known hyperthermophilic microbes were absent from the entire core. Overall our results shed new light on the distribution and composition of the boundary microbial community close to the high-temperature limit for habitability in the subseafloor environment of a hydrothermal field.
Similar content being viewed by others


Introduction
Deep-sea hydrothermal vents are extreme environments, especially with respect to the high temperatures, which create the limits of life on Earth. Several studies have further addressed the potential microbial habitats beneath the seafloor hydrothermal vents, the so-called ‘sub-vent biosphere’, and have indicated the possible occurrence of functionally active and metabolically diverse (hyper-)thermophilic microbial communities associated with shallow subseafloor hydrothermal fluids and mineral deposits (Deming and Baross, 1993; Delaney et al., 1998; Summit and Baross, 1998; Huber et al., 2002, 2003). The compositions and functions of the sub-vent microbial communities have been inferred from culture-dependent and culture-independent analyses of microbial communities in (i) in situ growth chambers placed in hydrothermal fluid flows (Karl et al., 1988; Reysenbach et al., 2000; Corre et al., 2001; Takai et al., 2004, ii) crustal fluids collected directly from the shallow subseafloor environments (~10 m below the seafloor [mbsf]) via seafloor drilling or probe insertion (Cowen et al., 2003; Higashi et al., 2004; Huber et al., 2006; Kato et al., 2009; Orcutt et al., 2011, iii) hydrothermal sediments (Teske et al., 2002; Dhillon et al., 2005; Nunoura et al., 2010; Teske et al., 2014; Dowell et al., 2016; Teske et al., 2016) and (iv) chimney structures of active hydrothermal vents (Takai et al., 2001; Schrenk et al., 2003; Nakagawa et al., 2005). These studies found that mesophilic, thermophilic and hyperthermophilic members of Epsilonproteobacteria, Gammaproteobacteria, Aquificales, Thermococcales and Methanococales were the potentially predominant microbial components in the sub-vent biosphere. In addition, previous studies associated with international scientific ocean drilling projects have also indicated the existence of microbial cells in deep subsurface sedimentary and rocky habitats in the hydrothermal vent systems on the Juan de Fuca Ridge (Ocean Drilling Program (ODP) Leg 139 (Cragg and Parkes, 1994) and 169 (Cragg et al., 2000; Summit et al., 2000) and Integrated Ocean Drilling Program (IODP) Expedition 301 (Lever et al., 2013)) and the Manus Basin (ODP Leg 193 (Kimura et al., 2003)). However, these microbial explorations associated with scientific ocean drilling projects have not successfully provided data on the compositions and functions of potential subseafloor microbial communities. In particular, it is unclear how local hydrothermal flow may influence the uneven distribution of sub-vent microbial community. Complex subsurface hydrothermal flow may limit or stimulate the sub-vent microbial activities.
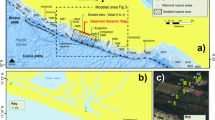
The IODP Expedition 331 by the D/V Chikyu (Takai et al., 2011) provided a good opportunity for direct investigation of the sub-vent biosphere at the Iheya North hydrothermal field in the Mid-Okinawa Trough. The Iheya North field is located in a continental-margin backarc basin, and the hydrothermal activity is highly influenced by trough-filling terrigenous sediment and knoll-covering volcaniclastic deposits (Kawagucci et al., 2011). The subseafloor geochemical and microbiological processes associated with buried organic matter and hydrothermal circulation produce unique hydrothermal fluid compositions that are enriched with high concentrations of methane and ammonia (Sakai et al., 1990; Kawagucci et al., 2011). During IODP Exp. 331, we conducted drilling and coring operations at five sites, located at 0 m (C0016), 100 m (C0013), 450 m (C0014) and 1550 m (C0017) east and 600 m (C0015) northwest of the most active hydrothermal mound (Takai et al., 2011). Previously, we have reported downhole changes in the microbial community corresponding to variations in the subseafloor hydrogeologic and lithostratigraphic structures that affected the recharging seawater and hydrothermal fluid input at Site C0017 (Yanagawa et al., 2014). That study obtained the 16S rRNA gene sequences of putative thermophiles only from the deepest sediments at 141 mbsf, where the in situ temperature was estimated to be approximately 90 °C, lower than the previously reported upper growth temperature limit of hyperthermophiles (122 °C) (Takai et al., 2008). The results suggested that the deepest sediment at Site C0017 is inhabited by potential thermophilic populations and that the habitable limit of the subseafloor microbial community lies at a depth greater than that of the bottom of the drilled hole (Yanagawa et al., 2014).
In addition to Site C0017, this study investigated the subseafloor environments at two additional drilling sites (Sites C0013 and C0014) that have higher thermal gradients and are closer (100 and 450 m, respectively) to the vigorous hydrothermal vents. We chose these vents to clarify the distribution, composition and function of microbial communities occurring adjacent to the high-temperature biosphere limit associated with high-temperature hydrothermal fluids. The drilling operations at these sites successfully penetrated the subseafloor hydrothermal fluid reservoirs and created artificial hydrothermal fluid vents with temperatures of up to 311 °C (Kawagucci et al., 2013a). Hence, the recovered core samples were potentially exposed to the in situ temperature range of 4 to >300 °C and provide important insights into the boundary microbial community and the limits of the biosphere.
Materials and methods
Sampling sites and sample collection
Subseafloor drilling and coring operations were conducted at the Iheya North hydrothermal field in the Mid-Okinawa Trough during IODP Expedition 331 (Takai et al., 2011). Coring and in situ temperature measurements were conducted at IODP Site C0013 and Site C0014 as previously described (Takai et al., 2011). Sediment samples for geochemical and microbiological analyses were collected as previously described (Yanagawa et al., 2013b, 2014). Details on the sample collections are provided in Supplementary Information Materials and Methods.
Geochemical analyses
The carbon and hydrogen isotopic compositions of methane were determined via continuous-flow isotope ratio mass spectrometry as previously described (Umezawa et al., 2009; Kawagucci et al., 2013a). The oxygen isotopic compositions of hydrothermally altered mud were determined as previously described (Miyoshi, 2013). The concentration and isotopic composition of dissolved inorganic carbon (DIC) was determined as previously described (Miyajima et al., 1995; Toki et al., 2004; Noguchi et al., 2013). Details are provided in Supplementary Information Materials and methods.
Molecular analyses of 16S rRNA genes and functional genes
DNA was extracted from core subsamples using the PowerMAX Soil DNA isolation kit (MoBio Laboratories, Carlsbad, CA, USA). Before the physical cell disruption, the samples were incubated at 65 °C for 5 min. Then, mechanical shaking was performed for 10 min with a ShakeMaster (BioMedical Science, Tokyo, Japan). Other subsequent steps were performed according to the manufacturer's protocol. The extracted DNA was stored at −80 °C.
Quantitative real-time PCR (Q-PCR) was performed for prokaryotic and archaeal 16S rRNA genes and the functional genes of dsrA, aprA and mcrA as previously described (Nunoura et al., 2008; Blazejak and Schippers, 2011; Yoshida-Takashima et al., 2012; Breuker et al., 2013). The primers, probes and amplification conditions used in this study are summarized in Supplementary Table S2.
For the clone analysis of 16S rRNA genes, the gene fragments were amplified by PCR using a universal and archaea-specific primer set (Nunoura et al., 2012). The mcrA gene fragments were also amplified using specific primers. Details for primers and PCR amplification conditions are described in Supplementary Table S2. The amplified 16S rRNA and mcrA genes were cloned, sequenced and aligned as previously described (Yanagawa et al., 2013b). The phylogenetic affiliations were identified using the SILVA SSU Ref 111 Database (Ludwig et al., 2004) and phylogenetic trees were constructed. A detailed description is provided in Supplementary Information Materials and Methods. Sequences have been deposited in the GenBank database under accession numbers: AB824899-AB825952 for 16S rRNA gene sequences and LC061224-LC061266 for mcrA gene sequences.
Cultivation tests
Serial dilution cultivations for anaerobic heterotrophs, methanogens, and sulfate reducers were performed to quantify the abundance of populations that can be cultivated in the laboratory using sediment slurries from a variety of depths. The cultivated population abundance of aerobic heterotrophs was quantified by colony-forming units on marine agar 2216 (BD). Details for cultivation conditions are provided in Supplementary Information Materials and methods and Supplementary Table S3.
Microbial activity measurements
The potential rates of methane production and oxidation and acetate oxidation were determined through radioisotope-tracer incubation experiments (Tasumi et al., 2015). The sediment slurry samples were incubated with appropriate radiotracers (14CH4, H14CO3− and [2-14C] acetate) for 1 month at 30 and 60 °C based on the in situ temperatures in Supplementary Figure S1. The radioactivity of a portion of the reaction products in the headspace was measured using a gas chromatograph (Shimadzu GC-2014, Shimadzu, Kyoto, Japan) and a high-sensitivity radioactivity detector RAGA Star (Raytest, Straubenhart, Germany), as shown in Supplementary Information Materials and methods. The potential activity was calculated based on the proportion of radioactive 14C product to total radioactive substrate.
Results and Discussion
Temperature in the sub-vent environment
Sediment core samples analyzed in this study were obtained from two holes at IODP Site C0013 (Holes C0013B and C0013D) and four holes at Site C0014 (Holes C0014B, C0014D, C0014E and C0014G). The depths of the deepest holes were 35 and 137 mbsf at Site C0013 (Hole C0013D) and Site C0014 (Hole C0014G), respectively. Although in situ temperatures were not measured during the drilling at Site C0013, it could be estimated from the fact that most of the plastic core liners melted several meters below the seafloor (Yanagawa et al., 2013b). The melting point of the plastic is >70 °C. The subseafloor environment at Site C0014, at which most of the deep coring operations were conducted by using aluminum core liners, exhibited a steep temperature gradient of 3 °C m−1 based on the temperatures measured during the drilling operation. At this site, the temperatures increased from 4.5 °C at the seafloor to >210 °C at 50 mbsf (Takai et al., 2011; Supplementary Figure S1).
Hydrothermally altered mud with elemental sulfur and sulfide grit was found in shallow sediments at 0.8 and 4.5 mbsf in Holes C0013B and C0013D, respectively (Miyoshi et al., 2015). The clay minerals in the hydrothermal mud yielded oxygen isotopic values of +8.0 and +9.6‰ (Miyoshi, 2013). These values indicated that the clay minerals formed at >138 °C (Supplementary Table S1). At Site C0014, the shallow sediments were not affected by hydrothermal alteration at depths above ~12 mbsf, whereas the deeper >12 mbsf sediments were hydrothermally altered (Miyoshi et al., 2015). The oxygen isotopic values of the hydrothermally altered clays at 12.8 and 15.1 mbsf in Hole C0014B were +8.7 and +12.5‰, respectively (Miyoshi, 2013). This indicated that the clay formation temperatures were >106 °C at Site C0014 (Supplementary Table S1; Miyoshi, 2013). The geochemical thermometer estimate based on the oxygen isotopic compositions of hydrothermally altered clay minerals suggests that at some point in time, even the shallow sediments at Sites C0013 and C0014 have experienced high-temperature conditions that are close to the known temperature limit of life at 122 °C (Takai et al., 2008). Indeed, any of the microbial cell observations, metabolic activity measurements, cultivation tests and prokaryotic 16S rRNA gene quantifications described below provided positive signatures for the existence of microbial populations only in the near-surface sediments at Site C0013. Thus, most of the results and discussion in the following sections are described with a focus on Site C0014.
A rapid increase in the pore-water potassium (K) concentration was observed just below the seafloor at 12.1 mbsf in Hole C0014B, 11.5 mbsf in Hole C0014D and 17.7 mbsf in Hole C0014G (Supplementary Figure S2). These discontinuities in K concentrations could be explained by the possible occurrence of impermeable layers in the subseafloor environment (Figure 1) and hydrothermal fluid flows just below the layers. The distribution pattern of the hydrothermally altered mud supports this interpretation (Supplementary Table S1; Miyoshi et al., 2015). In addition, because potassium was enriched in the endmember 310 °C hydrothermal fluids (80 mm) but low in the ambient 4 °C seawater (10 mm; Kawagucci et al., 2011), the K concentration is an indicator of the degree of mixing between hydrothermal fluids and infiltrated seawater and/or ambient pore-water. Although the magnesium concentration is also an excellent indicator of mixing between ambient seawater and hydrothermal fluids, the pore-water magnesium concentration could be affected by not only mixing but also association and dissociation processes related to hydrothermally altered minerals (Takai et al., 2011; Miyoshi et al., 2015). Thus, in this study, the pore-water K concentration is likely more reliable than the magnesium concentration. Assuming a bimodal mixing between endmember hydrothermal fluid and seawater without conductive cooling, K concentrations lower than 37 mm correspond to the temperature range of sediments lower than 122 °C, the potential habitable temperature range of previously cultivated microorganisms. In other words, K concentrations higher than 37 mm indicate that the sediments are exposed to high temperatures exceeding the potentially habitable temperature range.

Pore-water concentrations of methane and sulfate (Takai et al., 2011) (a), the carbon and hydrogen isotopic composition of methane (b), the numbers of 16S rRNA and functional genes (c and d) and heterotrophic activity (e) in the subseafloor core samples at IODP Site C0014. Q-PCR was used to quantify numbers of 16S rRNA genes of prokaryotes (black circles) and archaea (red diamonds), and functional genes of mcrA (purple diamonds), dsrA (green triangles) and aprA (orange squares). The total cell counts (gray crosses) were originally reported by Takai et al. (2011). Heterotrophic activity was determined based on the potential activity of the anaerobic oxidation of acetate. The incubations were conducted at 30 and 60 °C based on the in situ temperatures in Supplementary Figure S1. The samples incubated at 60 °C were indicated with parentheses, next to each symbol. Values below the detection limit for the Q-PCR data and heterotrophic activity are plotted as open symbols on the left axes. The red-shaded layers indicate the depth range of the limits for microbes based on Q-PCR data and heterotrophic activity. These depth ranges correspond to the depths of 14.3–17.2 mbsf in Hole C0014B, 10.2–11.4 mbsf in Hole C0014D and 15.8–17.6 mbsf in Hole C0014G. The possible impermeable layers in Supplementary Figure S2 are indicated by the blue dashed lines.
Geochemical evidence of hydrothermal fluid input and methane oxidation
The total alkalinity and DIC concentrations featured significant peaks at shallow depths in Holes C0014B and C0014D (Supplementary Figure S2). On the basis of the stable carbon isotope compositions, this enrichment was probably caused by CO2 in the endmember hydrothermal fluids. However, the DIC concentration peaks did not match the vertical K concentration profiles in each of the holes even though both components could have been provided by the hydrothermal fluids (Supplementary Figure S2). The different profiles for these two components (DIC and K) indicated that at least two different hydrothermal fluid sources fed the different lithostratigraphic horizons, such as the depth zones of 1–8.4 mbsf and >12.1 mbsf in the case of Hole C0014B. The hydrothermal fluid input in the shallower sediments (1–8.4 mbsf in Hole C0014B and 0–2.2 mbsf at C0014D) consisted primarily of low-temperature liquid CO2 and/or enriched gas components, whereas the input in the deeper zones (>12.1, >11.5 and >17.7 mbsf in Holes C0014B, C0014DE and C0014G, respectively) was derived directly from high-temperature vapor-lost fluid. Both of these fluids could result from the subseafloor phase separation and phase-partitioning processes of hydrothermal fluids (Von Damm and Bischoff, 1987).
The methane and sulfate concentrations exhibited more complex vertical variations than the DIC and K concentrations (Figure 1). The sulfate concentration exceeding the seawater level at Holes C0013B and D (Supplementary Figure S2) was probably due to the dissolution of anhydrite with decreasing temperature during the core recovery and sampling processes (Takai et al., 2011). In contrast, sulfate depletion and methane enrichment (relative to seawater levels) were apparent even within the shallow depths for each hole at Site C0014 (Figure 1a). Interestingly, a clear inverse relationship existed between methane and sulfate above 14 mbsf in Hole C0014B. This indicated the lateral flows of methane-rich hydrothermal fluids in the sulfate-rich subseafloor environment. In addition to the lithostratigraphically controlled hydrothermal fluid inputs, the δ13CCH4 and δDCH4 profiles suggested the contribution of microbial community functions. The patterns in the δ13CCH4 values along the vertical extent of the core were synchronized with δDCH4 values, and isotopically positive shifts in both the δ13C and δD values of methane were observed only within the shallow low-temperature zones (7.8–10.7 mbsf in Hole C0014B and <15.9 mbsf in Hole C0014G), where the methane concentration was extremely low (Figures 1a and 2). These isotopically heavier shifts in methane were not directly derived from the 13C- and D-depleted methane in the high-temperature hydrothermal fluids (Kawagucci, 2015). The concurrent isotopic changes in methane (ΔδDCH4/Δδ13CCH4) in the shallow low-temperature zones fell into a range bracketed by slopes of 7–19 (Figure 2), which have been reported to be representative values for microbial methane oxidation (Alperin et al., 1988; Kessler et al., 2006; Holler et al., 2009; Feisthauer et al., 2011).

Cross plots of δ13CCH4 and δDCH4 values. The open star represents the previously reported high-temperature hydrothermal fluid value (Kawagucci et al., 2011). Open and filled symbols represent samples obtained from shallow (low-temperature) and deep (high-temperature) zones in each hole, respectively (criteria of the depth are shown in Figure 1). Circles, squares and diamonds represent Holes C0014B, C0014DE and C0014G, respectively. Representative values for thermogenic and biogenic methane (Kawagucci et al., 2013b) are shown by a shaded area and an arrow, respectively. The diagonal lines, with slopes of 7 and 19, represent the lowest and highest values of the co-variation exhibited in microbial methane oxidation.
The vertical patterns of the geochemical signals differed among the holes (Figures 1a and b; Supplementary Figure S2). The spatial heterogeneity in the subseafloor pore-water geochemistry was likely affected by the local input of hydrothermal fluids, and the observed patterns, particularly in Hole C0014B, indicated vertical differences in fluid chemistry due to different sources of laterally flowing hydrothermal fluids and their mixing behavior with infiltrated seawater and ambient pore-water. In combination with the complex variations in the lithostratigraphic and physical properties of the core samples, the phase separation, flow patterns and mixing behavior of subseafloor hydrothermal fluids are hypothesized to be controlled by the lithostratigraphy, such as fresh hemipelagic sediments, porous pumiceous deposits and hard impermeable layers (Takai et al., 2011).
Abundance estimation of microbial cellular and rRNA gene populations
The microbial cell abundance at Site C0014 decreased markedly with depth from approximately 1 × 108 cells per ml of sediment just below the seafloor to less than the detection limit of approximately 6.5 × 105 cells per ml of sediment (Takai et al., 2011; Figure 1c). A Q-PCR analysis for whole prokaryotic and archaeal 16S rRNA genes suggested that microbial populations were present only in the shallow sediments above 14.3, 10.2 and 15.8 mbsf in Holes C0014B, C0014E and C0014G, respectively (Figure 1c). The abundance of the whole prokaryotic 16S rRNA genes with depth showed similar values and profiles to the microbial cell abundance in all of the holes. However, several shallow sediment depths in Holes C0014B and C0014G exhibited large data gaps between the microbial cell abundance and the whole prokaryotic 16S rRNA gene abundance due to the relatively higher detection limit of the microbial cell count, as shown in Figure 1c, or due to the technical limitations of the Q-PCR (Hoshino and Inagaki, 2012; Lloyd et al., 2013; Morono et al., 2014). The relative abundance of archaeal 16S rRNA genes increased with depth and represented approximately half of the whole prokaryotic 16S rRNA gene assemblages in the deepest zones where Q-PCR could detect 16S rRNA genes (Supplementary Figure S3).
Cultivation and activity measurements for heterotrophs
Successful enrichments of heterotrophs were obtained from most of the shallow sediment samples from Hole C0014B at a temperature of 30 °C, and subsequent quantitative cultivation analyses were conducted on these samples. Aerobic heterotrophs, anaerobic heterotrophs and sulfate reducers were detected at depths above 8.5, 12.2 and 3.6 mbsf, respectively (Supplementary Figure S4). In contrast, 60 °C incubations and onboard cultivation experiments for (hyper-)thermophilic heterotrophs and chemolithoautotrophs did not yield any cultivable population at any depth (Takai et al., 2011). The cultivated populations ranged up to 9.6 × 105 CFU g−1 sediment for aerobic heterotrophs, 5.5 × 107 cells g−1 sediment for anaerobic heterotrophs and 5.5 × 102 cells g−1 sediment for sulfate reducers. The 16S rRNA gene sequences of the isolated aerobic heterotrophs, anaerobic heterotrophs and sulfate reducers were highly similar to those of Geofilum rubicundum (100% similarity) [NR_112717], Clostridium sp. S710(0)-1 (98% similarity) [GU136592] and Desulfomicrobium norvegicum (100% similarity) [NR_025407], respectively (Supplementary Table S3). Potential anaerobic heterotrophic activities were also detected in the sediment samples from which the anaerobic heterotrophs were successfully cultivated (Figure 1e). The activity, defined as the oxidation of the methyl group of 14C-labeled acetate to 14CO2, gradually decreased with increasing sediment depth. Notably, all of the estimations of cultivation-dependent viable heterotrophic populations, 14C-tracer heterotrophic activities and prokaryotic 16S rRNA gene quantification provided independent detectable signatures for the occurrence of a microbial community in the same depth zone of the subseafloor environment (Figures 1c and e; Supplementary Figure S4).
16S rRNA gene community structures
The primer set of Uni530F/Uni907R for the universal prokaryotic 16S rRNA gene (Nunoura et al., 2012) was applied to DNA assemblages extracted from the sediment samples. Potentially indigenous 16S rRNA gene communities were obtained from the sediment samples from depths above 8.5, 10.2 and 15.8 mbsf in Holes C0014B, C0014D and C0014G, respectively (Figure 3a). None of the 16S rRNA genes were amplified from environmental DNA assemblages below these depths. The 16S rRNA gene phylotype compositions changed significantly with transitions in the geochemical and lithostratigraphical environments and/or the elevated temperatures. Typical uncultivated microbial members in the marine sedimentary environment, such as Deltaproteobacteria, Chloroflexi, JS1 group in Candidatus (Ca.) Atribacteria, Deep-sea Hydrothermal Vent Euryarchaeotic Group 6 (DHVEG-6) and Miscellaneous Crenarchaeotic Group (MCG), were detected in the 16S rRNA gene clone communities at all depths. Certain members closely related to potentially thermophilic bacterial taxa/divisions, such as Thermotogae, Thermodesulfobacteria and OP1 (Ca. Acetothermia), were detected as minor populations in certain deeper sections (Supplementary Data S1).

16S rRNA gene phylotype compositions in the sediments from Site C0014, displayed (a) with respect to sediment depth using a universal primer set (Uni530F-907R) and (b) as pie diagrams at specific depths using an archaea-specific primer set (Arc530F-Arc958R). Archaeal 16S rRNA gene amplicons were obtained from three samples, which were not amplified with Uni530F-907R (shown as ‘ND’ in the black column). The red dotted lines denote the cutoff between Bacteria and Archaea. The numbers in parentheses indicate the number of clones. The red-shaded layers represent the possible depth limit of active subseafloor microbes, as determined in Figure 1.
Members of the HotSeep-1 group composed 60–100% of the total number of deltaproteobacterial 16S rRNA gene sequences from the deeper sediment samples at 8.5 mbsf in Hole C0014B, 10.2 mbsf in Hole C0014D and 15.8 mbsf in Hole C0014G (Supplementary Data S1). The HotSeep-1 group was previously detected in hydrothermal sediments in the Guaymas Basin (Teske et al., 2002; Kniemeyer et al., 2007; Dowell et al., 2016) and in an enrichment culture of anaerobic methane oxidizers under high-temperature conditions (Holler et al., 2011). Hence, the HotSeep-1 group is thought to be responsible for high-temperature sulfate reduction, coupling with the anaerobic oxidation of methane (AOM) by a deeply branching, putatively thermophilic group of methanotrophic archaea (ANME-1-Guaymas I; Holler et al., 2011). Although the sequences related to anaerobic methanotrophs (ANMEs) were found in certain depth horizons, the members closely related to ANME-1-Guaymas I did not co-exist with the bacterial HotSeep-1 group. Notably, the ANME-1-Guaymas I phylotypes were detected only at 14.3 mbsf in Hole C0014B, representing only 2% of the archaeal clone library (Supplementary Data S1).
Archaeal 16S rRNA gene amplicons were obtained from samples from the deepest zones at Site C0014, at which many positive signals for the existence of microbial communities were detected. These archaeal 16S rRNA gene amplicons were detected not by using a universal primer set but by using the archaea-specific primer set Arch530F/Arch958R (Figure 3b). The archaeal 16S rRNA gene communities in these sediments were dominated (42–97% of total archaeal sequences) by the Hot Water Crenarchaeotic Group IV (HWCGIV; also known as UCII or a subcluster of the Terrestrial Hot Spring Crenarchaeotic Group [THSCG]; Supplementary Data S1). They were also detected as the predominant population (99% of the total archaeal sequences) in the near-surface sediment at 1 mbsf in Hole C0013B, which is the only sample containing detectable 16S rRNA genes at Site C0013 (data not shown). The HWCGIV sequences have previously been reported in deep-sea hydrothermal vent habitats (Schrenk et al., 2003; Nunoura et al., 2010; Yoshida-Takashima et al., 2012) and in a high-temperature zone at 141.1 mbsf at Site C0017 (Yanagawa et al., 2014).
Microbial functions of methanogenesis and AOM
The metabolic activities of hydrogenotrophic methanogenesis, acetoclastic methanogenesis and AOM were traced using radioisotope-labeled substrates at in situ temperatures. Hydrogenotrophic and acetoclastic methanogenic activities were detected at certain depths in the relatively shallow zones at Site C0014, whereas AOM activity occurred widely in the subseafloor environment, from which positive signals of the existence of microbial communities were obtained (Figure 4). Although the hydrogenotrophic methanogenesis activity outcompeted the AOM activity in the shallower zone and the AOM activity dominated in the deeper zone, the opposing microbial processes of methane production and consumption occurred at the same depths (0.3, 3.6 and 5.0 mbsf in Hole C0014B, 0.2 mbsf in Hole C0014D and 0.3, 4.1 and 7.8 mbsf in Hole C0014G). In addition, relatively high AOM activities and extremely low pore-water sulfate concentrations were observed at the approximate depths of 3.6, 8.6 and 15.8 mbsf in Holes C0014B, C0014D and C0014G, respectively (Figure 1a).

Site C0014 depth profiles of the potential activity of AOM, hydrogenotrophic methanogenesis and acetoclastic methanogenesis (from left to right). The phylogenetic affiliation of mcrA genes is shown in the rightmost column. Open circles on the y-axes denote analyses below the detection limit. The red-shaded layers represent the possible depth limit of active subseafloor microbes, as determined in Figure 1.
Phylogenetic diversity and the abundance of functional genes related to methanogenesis and AOM were also characterized via both Q-PCR for genes associated with methyl coenzyme M reductase (mcrA), dissimilatory sulfite reductase (dsrA) and adenosine 5′-phosphosulfate reductase (aprA) and clone analysis for mcrA (Figures 1d and 4). The Q-PCR data indicated that the mcrA, dsrA and aprA genes were widely distributed in most of the shallow sediments that yielded many positive signals for the existence of microbial communities, and the abundances decreased with increasing depth in the sediment. This pattern also indicated that the complex biogeochemical processes associated with the microbial methane- and sulfate-related metabolisms co-occur in the sub-vent biosphere at Site C0014. The abundances of these functional genes were one to two orders of magnitude lower than those of the prokaryotic 16S rRNA genes in the same samples.
On the basis of the sequences of the mcrA genes obtained from the clone analysis, the mcrA genes were conventionally classified into the types derived from potential methanogenic and methanotrophic populations. The methanogenic type of mcrA genes were obtained only from the uppermost sediment at 0.3 mbsf in Hole C0014B (Figure 4) and were phylogenetically related to the mcrA genes of Methanococcoides (Supplementary Figure S5). The mcrA group a-b genes, which were derived from the ANME-1 (Knittel and Boetius, 2009), were most frequently obtained throughout the entire depth of the potential microbial habitable zone at Site C0014. In contrast, most of the mcrA gene sequences of groups c-d, e and f, hosted by ANME-2a, -2c and -3 (Knittel and Boetius, 2009), respectively, were found in the shallower depths of sediments (Figure 4). Molecular ecological studies have indicated that the ANME-1 populations occur in deeper, more reductive and more sulfate-depleted habitats than the ANME-2 populations (Knittel et al., 2005; Krüger et al., 2008; Nunoura et al., 2008; Rossel et al., 2011; Yanagawa et al., 2011). Furthermore, the potentially thermophilic ANME-1 group has been recognized as a key component in certain hydrothermal ecosystems, such as Guaymas Basin and Juan de Fuca Ridge (Biddle et al., 2012; Lever et al., 2013; Merkel et al., 2013). This mcrA group of thermophilic ANME-1 is defined as Hydrothermal ANME-1 Cluster II (Lever et al., 2013) (alternatively classified as mcrA-Guaymas (Biddle et al., 2012) or ANME-1GBa (Merkel et al., 2013)). The optimal growth temperatures of the thermophilic ANME-1 in the Guaymas site have been estimated to be above 70 °C (Merkel et al., 2013). We detected the same group of mcrA gene sequences at 6.7 mbsf in Hole C0014D and 15.8 mbsf in Hole C0014G (Figure 4; Supplementary Figure S5). The bacterial 16S rRNA gene sequences of potential thermophiles (Thermotogae and OP1) were also detected at these sediment depths (Supplementary Data S1). Thus, the possible host archaeal populations of the Hydrothermal ANME-1 Cluster II mcrA genes may be thermophilic. Indeed, the radioisotope-tracer AOM activity measurements showed relatively high activity, 3.1 pmol cm−3 d−1, at 60 °C at 15.8 mbsf in Hole C0014G (Figure 4).
Synthesis and interpretation
Development of methane-consuming microbial communities
Previous studies of microbial communities associated with the hydrothermal vent fluids and chimney deposits in the Iheya North hydrothermal field have hypothesized the existence of active sub-vent microbial communities that are potentially dominated by hyperthermophilic Thermococcales and chemolithoautotrophs with H2- and/or sulfur-compound metabolisms. These metabolisms were extrapolated from the variability in the microbial communities in the most interior and/or reductive seafloor habitats that are highly affected by the hydrothermal fluid input (Nakagawa et al., 2005; Takai et al., 2006). However, several thermodynamic estimates of chemolithotrophic microbial community development in hydrothermal-mixing zones have suggested that the populations that couple anaerobic methanotrophy with sulfate reduction would energetically dominate the microbial communities in the sediment-hosted seafloor and subseafloor habitats that are highly affected by hydrothermal fluid inputs (Takai and Nakamura, 2011; Nakamura and Takai, 2014; Takai et al., 2014). In this study, all of the pore-water geochemical analyses, metabolic activity measurements and cellular and molecular microbial community analyses indicated the occurrence of functionally active microbial communities dominated by AOM populations in the relatively shallow subseafloor habitats down to 15.8 mbsf in Hole C0014G. Several studies have examined the abundance, phylogenetic diversity and function of AOM populations associated with seafloor hydrothermal activity in the Guaymas Basin and Yonaguni Knoll IV fields (Teske et al., 2002; Nunoura et al., 2010; Yanagawa et al., 2013a; Dowell et al., 2016). Because these investigations have focused on the shallow sediments just beneath the seafloor, where the diffusive mixing of hydrothermal fluids and seawater likely characterizes the geochemical environments, there remains a lack of knowledge on the subseafloor AOM communities associated with hydrogeologically controlled advection and the partitioning and mixing processes of hydrothermal fluids and infiltrated seawater near deep-sea vents.
The vertical profiles of pore-water methane and sulfate concentrations (Figure 1a) and stable isotopic values of δ13CCH4 and δDCH4 (Figure 2) indicated the abundant occurrence of potentially sulfate-reducing AOM functions in several specific horizons within the microbially habitable subseafloor environment (for example, depths of 0–11.7 mbsf in Hole C0014B and 0–10.2 and 0–15.8 mbsf in Hole C0014G). In addition, previous study of significant 34S enrichment in pore-water sulfate strongly suggested the presence of microbial sulfate reduction at several depths within the microbially habitable terrain, that is, ~5 mbsf in Hole C0014B and 16.1 mbsf in Hole C0014G (Aoyama et al., 2014). These compositional and isotopic profiles of pore-water methane and sulfate are largely dependent on the mass balance of hydrothermal fluid and seawater inputs and in situ microbial consumption. Thus, the geochemically identified peaks and valleys in the potential microbial methane oxidation and sulfate reduction are not necessarily equivalent to the abundance of in situ microbial functions. Indeed, the potential in situ microbial activity profile of AOM, as determined via radioisotope-tracer experiments, did not match the geochemically identified peaks and valleys of microbial methane oxidation (Figure 4), and the mcrA and dsrA gene abundances gradually decreased with increasing depth (Figure 1d). However, overall, the pore-water geochemistry, the in situ metabolic activities and the 16S rRNA and functional gene distributions indicated a significant association between subseafloor AOM activity and sulfate reduction. On the other hand, previous studies have suggested that microbial AOM activity might be coupled to the reduction of iron (hydr)oxide minerals but not sulfate reduction in metalliferous hydrothermal sediments (Wankel et al., 2012). The physical and chemical variations in the subseafloor environment resulting from advective hydrothermal fluid and seawater flows related to the lithostratigraphic and hydrogeologic conditions would create diverse biogeochemical processes.
Constraints on microbial community development in the sub-vent biosphere
All of the direct microscopic observations, PCR-based molecular analyses, cultivation tests and metabolic activity measurements indicated that functionally active, metabolically diverse microbial communities developed in the shallow zones of subseafloor sediments associated with hydrothermal fluid flows. The positive signals of the existence of microbial communities were obtained from the sediments down to 14.3, 10.2 and 15.8 mbsf in Holes C0014B, C0014D and C0014G, respectively. Microbial populations were not detected in the deeper, high-temperature hydrothermal fluid regimes, due to the limit of microbial habitability and/or the methodological detection limit in this study. The Q-PCR and clone library analyses for 16S rRNA genes showed that the abundance of archaeal 16S rRNA gene phylotypes increased with depth, whereas bacterial phylotype populations dominated the microbial communities at shallower depths (Supplementary Figure S3). However, neither the culture-dependent nor the culture-independent analyses detected the existence of certain previously cultivated hyperthermophilic populations, such as Thermococcales, Methanococcales, Archaeoglobales and members of Crenarchaota, even though such hyperthermophilic archaeal populations are known to dominate the microbial communities in other high-temperature hydrothermal fluid and chimney habitats (Nakagawa et al., 2005; Takai et al., 2006). One conceivable explanation for this may relate to unstable temperature conditions for their growth due to fluctuating hydrothermal fluids, as described below. In the 16S rRNA gene clone libraries obtained from the deepest sediments of the microbially habitable terrain in the subseafloor environment, putative thermophilic phylotypes, such as members of HWCGIV, thermophilic ANME-1, OP1 and Thermotogae, were found (Figure 3; Supplementary Data S1). According to the molecular thermometer calculation based on the GC content of the 16S rRNA gene sequences (Kimura et al., 2010), the potential growth temperature range was estimated to be the highest (50 –76 °C) for members of HWCGIV among the putative thermophilic populations.
The relationship between microbial habitability and the in situ temperature of the subseafloor biosphere has rarely been investigated. Recently, an active subseafloor microbial community has been discovered in deeply buried terrigenous sediments with a temperature of <60 °C at 2458 mbsf (Inagaki et al., 2015). The low abundance of this community was attributed to the increase in energy used for the repair of essential biomolecules such as amino acid and DNA. The energetic costs of amino acid racemization and DNA depurination increase exponentially with temperature (Lever et al., 2015). However, the physical and chemical conditions that form the boundary between habitable and uninhabitable terrains have not been directly explored in subseafloor environments (Takai et al., 2014). Sub-vent microbial communities are believed to live near the upper temperature limits for life on this planet. However, based on the predicted thermal gradient from the temperatures measured during the drilling operation (Takai et al., 2011), the temperature near the detection limit of microbial populations, activities and molecules in Hole C0014B (14.3 mbsf) was estimated to be ca. 45 °C. In contrast, the geochemical thermometer estimate based on the oxygen isotopic compositions of hydrothermally altered clay minerals from 12.8–15.1 mbsf in Hole C0014B suggested that the boundary habitat experienced higher temperatures (>106 °C) (Supplementary Table S1; Supplementary Figure S1). The former temperature (ca. 45 °C) and latter (>106 °C) temperature are far below and similar to the upper temperature limit for microbial growth, respectively (Takai et al., 2008). Additionally, the molecular thermometer estimate based on the potentially most thermophilic 16S rRNA gene phylotype (HWCGIV) population at the depth limit yields a growth temperature range of 50–76 °C. It remains uncertain, which of the estimated temperatures most accurately represents a realistic in situ temperature range because the bottom temperature of present microbial habitable zone was not directly measured during the drilling operation, and the geochemical and molecular thermometer estimates do not necessarily reflect the current in situ temperature range. One plausible interpretation is that the deepest microbial habitat in Hole C0014B has been exposed to considerably high temperatures (>106 °C) in the past and presently experiences fluctuating temperatures that are induced by ever-varying degrees of mixing between the subseafloor high-temperature hydrothermal fluids, diffusive and/or advective seawater flows and conductive cooling. Similarly, the hydrothermally altered sediments that were observed in the shallow sediments at Site C0013 and the oxygen isotopic values of the clay minerals indicate formation temperatures of >138 °C (Miyoshi, 2013; Supplementary Table S1). The excess levels of pore-water sulfate concentrations (relative to the sulfate concentrations of seawater) in these sediments were attributed to anhydrite dissolution related to cooling during core recovery and sampling (Supplementary Figure S2). Because anhydrite is stable only under high temperatures (>150 °C) (Gieskes et al., 2002; Takai et al., 2011), the subseafloor sediments at Site C0013 that contain excess pore-water sulfate concentrations (relative to seawater) are likely exposed to high temperatures that exceed the microbially habitable range at the present. The microbial habitability in the sub-vent biosphere is probably highly constrained by the latest temperature history and/or the present temperature conditions induced by spatiotemporally variable high-temperature hydrothermal fluid input.
Furthermore, the physical properties of the sediments are another important factor constraining the microbially habitable terrain in the sub-vent environment. The possible impermeable layers, which are predicted from the discontinuities in the pore-water chemical compositions, serve as not only shields limiting vertical flow and diffusion of fluids but also as barriers limiting vertical migration of microbial cells. Interestingly, the potential limit of microbial community development was always located above or near the possible impermeable layers at Site C0014 (Figure 1). Fluctuations in temperature in the boundary habitat probably occasionally sterilize the microbial community via exposure to high-temperature hydrothermal fluids, and this is followed by a gradual return to microbially habitable temperatures. However, the existence of impermeable layers may contribute to the slow recolonization of the active microbial community after sterilization because the impermeable layer limits migration of viable populations from refugia. In addition, the impermeable layers may also serve as the possible microbial refugia from the lethal temperature fluctuations associated with the occasional exposure to high-temperature hydrothermal fluids. The slow but successful recolonization of the active microbial community after a temporary sterilization may be initiated from viable microbial cells disseminated throughout the safe interior of impermeable layers. Although the permeable and impermeable inter-layer sequences of cores are often difficult to recover in IODP-like drilling operations, these hypotheses of the limit and recolonization of the sub-vent biosphere should be clarified in future research.
References
Alperin MJ, Reeburgh WS, Whiticar MJ . (1988). Carbon and hydrogen fractionation resulting from anaerobic methane oxidation. Global Biogeochem Cycles 2: 279–288.
Aoyama S, Nishizawa M, Takai K, Ueno Y . (2014). Microbial sulfate reduction within the Iheya North subseafloor hydrothermal system constrained by quadruple sulfur isotopes. Earth Planet Sci Lett 398: 113–126.
Biddle JF, Cardman Z, Mendlovitz H, Albert DB, Lloyd KG, Boetius A et al. (2012). Anaerobic oxidation of methane at different temperature regimes in Guaymas Basin hydrothermal sediments. ISME J 6: 1018–1031.
Blazejak A, Schippers A . (2011). Real-time PCR quantification and diversity analysis of the functional genes aprA and dsrA of sulfate-reducing prokaryotes in marine sediments of the Peru continental margin and the Black Sea. Front Microbiol 2: 253.
Breuker A, Stadler S, Schippers A . (2013). Microbial community analysis of deeply buried marine sediments of the New Jersey shallow shelf (IODP Expedition 313). FEMS Microbiol Ecol 85: 578–592.
Corre E, Reysenbach A-L, Prieur D . (2001). ɛ-proteobacterial diversity from a deep-sea hydrothermal vent on the Mid-Atlantic ridge. FEMS Microbiol Lett 205: 329–335.
Cowen JP, Giovannoni SJ, Kenig F, Johnson HP, Butterfield D, Rappé MS et al. (2003). Fluids from aging ocean crust that support microbial life. Science 299: 120–123.
Cragg BA, Parkes RJ . (1994). Bacterial profiles in hydrothermally active deep sediment layers from Middle Valley (NE Pacific) sites 857 and 858. Proc Ocean Drill Prog Sci Results 139: 509–516.
Cragg BA, Summit M, Parkes RJ . (2000). Bacterial profiles in a sulfide mound (site 1035) and an area of active fluid venting (site 1036) in hot hydrothermal sediments from Middle Valley (Northeast Pacific). Proc Ocean Drill Prog Sci Results 169: 1–18.
Delaney JR, Kelley DS, Lilley MD, Butterfield DA, Baross JA, Wilcock WSD et al. (1998). The quantum event of oceanic crustal accretion: impacts of diking at mid-ocean ridges. Science 281: 222–230.
Deming JW, Baross JA . (1993). Deep-sea smokers: Windows to a subsurface biosphere? Geochim Cosmochim Acta 57: 3219–3230.
Dhillon A, Lever M, Lloyd KG, Albert DB, Sogin ML, Teske A . (2005). Methanogen diversity evidenced by molecular characterization of methyl coenzyme M reductase A (mcrA genes in hydrothermal sediments of the Guaymas Basin. Appl Environ Microbiol 71: 4592–4601.
Dowell F, Cardman Z, Dasarathy S, Kellermann M, Lipp JS, Ruff SE et al. (2016). Microbial communities in methane- and short chain alkane-rich hydrothermal sediments of Guaymas Basin. Front Microbiol 7: 17.
Feisthauer S, Vogt C, Modrzynski J, Szlenkier M, Krüger M, Siegert M et al. (2011). Different types of methane monooxygenases produce similar carbon and hydrogen isotope fractionation patterns during methane oxidation. Geochim Cosmochim Acta 75: 1173–1184.
Gieskes JM, Simoneit BRT, Shanks Iii WC, Goodfellow WD, James RH, Baker PA et al. (2002). Geochemistry of fluid phases and sediments: relevance to hydrothermal circulation in Middle Valley, ODP Legs 139 and 169. Appl Geochem 17: 1381–1399.
Higashi Y, Sunamura M, Kitamura K, Nakamura K-i, Kurusu Y, Ishibashi J-i et al. (2004). Microbial diversity in hydrothermal surface to subsurface environments of Suiyo Seamount, Izu-Bonin Arc, using a catheter-type in situ growth chamber. FEMS Microbiol Ecol 47: 327–336.
Holler T, Wegener G, Knittel K, Boetius A, Brunner B, Kuypers MMM et al. (2009). Substantial 13C/12C and D/H fractionation during anaerobic oxidation of methane by marine consortia enriched in vitro. Environ Microbiol Rep 1: 370–376.
Holler T, Widdel F, Knittel K, Amann R, Kellermann MY, Hinrichs K-U et al. (2011). Thermophilic anaerobic oxidation of methane by marine microbial consortia. ISME J 5: 1946–1956.
Hoshino T, Inagaki F . (2012). Molecular quantification of environmental DNA using microfluidics and digital PCR. Syst Appl Microbiol 35: 390–395.
Huber JA, Butterfield DA, Baross JA . (2002). Temporal changes in archaeal diversity and chemistry in a mid-ocean ridge subseafloor habitat. Appl Environ Microbiol 68: 1585–1594.
Huber JA, Butterfield DA, Baross JA . (2003). Bacterial diversity in a subseafloor habitat following a deep-sea volcanic eruption. FEMS Microbiol Ecol 43: 393–409.
Huber JA, Johnson HP, Butterfield DA, Baross JA . (2006). Microbial life in ridge flank crustal fluids. Environ Microbiol 8: 88–99.
Inagaki F, Hinrichs K-U, Kubo Y, Bowles MW, Heuer VB, Hong W-L et al. (2015). Exploring deep microbial life in coal-bearing sediment down to ~2.5 km below the ocean floor. Science 349: 420–424.
Karl DM, Taylor GT, Novitsky JA, Jannasch HW, Wirsen CO, Pace NR et al. (1988). A microbiological study of Guaymas Basin high temperature hydrothermal vents. Deep Sea Res Part I Oceanogr Res Pap 35: 777–791.
Kato S, Yanagawa K, Sunamura M, Takano Y, Ishibashi J-i, Kakegawa T et al. (2009). Abundance of Zetaproteobacteria within crustal fluids in back-arc hydrothermal fields of the Southern Mariana Trough. Environ Microbiol 11: 3210–3222.
Kawagucci S . (2015).Fluid geochemistry of high-temperature hydrothermal fields in the Okinawa Trough. In: Ishibashi J-I, et al. (eds). Subseafloor Biosphere Linked to Hydrothermal Systems. Springer: Tokyo, Japan, pp 387–403.
Kawagucci S, Chiba H, Ishibashi J-i, Yamanaka T, Toki T, Muramatsu Y et al. (2011). Hydrothermal fluid geochemistry at the Iheya North field in the mid-Okinawa Trough:implication for origin of methane in subseafloor fluid circulation systems. Geochem J 45: 109–124.
Kawagucci S, Miyazaki J, Nakajima R, Nozaki T, Takaya Y, Kato Y et al. (2013a). Post-drilling changes in fluid discharge pattern, mineral deposition, and fluid chemistry in the Iheya North hydrothermal field, Okinawa Trough. Geochem Geophys Geosyst 14: 4774–4790.
Kawagucci S, Ueno Y, Takai K, Toki T, Ito M, Inoue K et al. (2013b). Geochemical origin of hydrothermal fluid methane in sediment-associated fields and its relevance to the geographical distribution of whole hydrothermal circulation. Chem Geol 339: 213–225.
Kessler JD, Reeburgh WS, Tyler SC . (2006). Controls on methane concentration and stable isotope (δ2H-CH4 and δ13C-CH4 distributions in the water columns of the Black Sea and Cariaco Basin. Global Biogeochem Cycles 20: GB4004.
Kimura H, Asada R, Masta A, Naganuma T . (2003). Distribution of microorganisms in the subsurface of the Manus Basin hydrothermal vent field in Papua New Guinea. Appl Environ Microbiol 69: 644–648.
Kimura H, Mori K, Tashiro T, Kato K, Yamanaka T, Ishibashi J-I et al. (2010). Culture-independent estimation of optimal and maximum growth temperatures of archaea in subsurface habitats based on the G+C content in 16S rRNA gene sequences. Geomicrobiol J 27: 114–122.
Kniemeyer O, Musat F, Sievert SM, Knittel K, Wilkes H, Blumenberg M et al. (2007). Anaerobic oxidation of short-chain hydrocarbons by marine sulphate-reducing bacteria. Nature 449: 898–901.
Knittel K, Boetius A . (2009). Anaerobic oxidation of methane: Progress with an unknown process. Annu Rev Microbiol 63: 311–334.
Knittel K, Lösekann T, Boetius A, Kort R, Amann R . (2005). Diversity and distribution of methanotrophic archaea at cold seeps. Appl Environ Microbiol 71: 467–479.
Krüger M, Blumenberg M, Kasten S, Wieland A, Kanel L, Klock JH et al. (2008). A novel, multi-layered methanotrophic microbial mat system growing on the sediment of the Black Sea. Environ Microbiol 10: 1934–1947.
Lever MA, Rogers KL, Lloyd KG, Overmann J, Schink B, Thauer RK et al. (2015). Life under extreme energy limitation: a synthesis of laboratory- and field-based investigations. FEMS Microbiol Rev 39: 688–728.
Lever MA, Rouxel O, Alt JC, Shimizu N, Ono S, Coggon RM et al. (2013). Evidence for microbial carbon and sulfur cycling in deeply buried ridge flank basalt. Science 339: 1305–1308.
Lloyd KG, May MK, Kevorkian RT, Steen AD . (2013). Meta-analysis of quantification methods shows that archaea and bacteria have similar abundances in the subseafloor. Appl Environ Microbiol 79: 7790–7799.
Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar et al. (2004). ARB: a software environment for sequence data. Nucleic Acids Res 32: 1363–1371.
Merkel AY, Huber JA, Chernyh NA, Bonch-Osmolovskaya EA, Lebedinsky AV . (2013). Detection of putatively thermophilic anaerobic methanotrophs in diffuse hydrothermal vent fluids. Appl Environ Microbiol 79: 915–923.
Miyajima T, Miyajima Y, Hanba YT, Yoshii K, Koitabashi T, Wada E . (1995). Determining the stable isotope ratio of total dissolved inorganic carbon in lake water by GC/C/IIRMS. Limnol Oceanogr 40: 994–1000.
Miyoshi Y, Ishibashi J-i, Shimada K, Inoue H, Uehara S, Tsukimura K . (2015). Clay minerals in an active hydrothermal field at Iheya-North-Knoll, Okinawa Trough. Resour Geol 65: 346–360.
Miyoshi Y . (2013).,Mineralogical and geochemical studies of hydrothermal clay minerals below the seafloor at active hydrothermal fields in the island arc and back-arc setting. PhD Dissertation, Kyushu University.
Morono Y, Terada T, Hoshino T, Inagaki F . (2014). Hot-alkaline DNA extraction method for deep-subseafloor archaeal communities. Appl Environ Microbiol 80: 1985–1994.
Nakagawa S, Takai K, Inagaki F, Chiba H, Ishibashi J-i, Kataoka S et al. (2005). Variability in microbial community and venting chemistry in a sediment-hosted backarc hydrothermal system: impacts of subseafloor phase-separation. FEMS Microbiol Ecol 54: 141–155.
Nakamura K, Takai K . (2014). Theoretical constraints of physical and chemical properties of hydrothermal fluids on variations in chemolithotrophic microbial communities in seafloor hydrothermal systems. Prog Earth Planet Sci 1: 1–24.
Noguchi T, Hatta M, Yamanaka T, Okamura K . (2013). Fast measurement of dissolved inorganic carbon concentration for small-volume interstitial water by acid extraction and nondispersive infrared gas analysis. Anal Sci 29: 9–13.
Nunoura T, Oida H, Miyazaki J, Miyashita A, Imachi H, Takai K . (2008). Quantification of mcrA by fluorescent PCR in methanogenic and methanotrophic microbial communities. FEMS Microbiol Ecol 64: 240–247.
Nunoura T, Oida H, Nakaseama M, Kosaka A, Ohkubo SB, Kikuchi T et al. (2010). Archaeal diversity and distribution along thermal and geochemical gradients in hydrothermal sediments at the Yonaguni Knoll IV hydrothermal field in the southern Okinawa Trough. Appl Environ Microbiol 76: 1198–1211.
Nunoura T, Takaki Y, Kazama H, Hirai M, Ashi J, Imachi H et al. (2012). Microbial diversity in deep-sea methane seep sediments presented by SSU rRNA gene tag sequencing. Microbes Environ 27: 382–390.
Orcutt BN, Bach W, Becker K, Fisher AT, Hentscher M, Toner BM et al. (2011). Colonization of subsurface microbial observatories deployed in young ocean crust. ISME J 5: 692–703.
Reysenbach A-L, Longnecker K, Kirshtein J . (2000). Novel bacterial and archaeal lineages from an in situ growth chamber deployed at a Mid-Atlantic Ridge hydrothermal vent. Appl Environ Microbiol 66: 3798–3806.
Rossel PE, Elvert M, Ramette A, Boetius A, Hinrichs K-U . (2011). Factors controlling the distribution of anaerobic methanotrophic communities in marine environments: evidence from intact polar membrane lipids. Geochim Cosmochim Acta 75: 164–184.
Sakai H, Gamo T, Kim E-S, Tsutsumi M, Tanaka T, Ishibashi J et al. (1990). Venting of carbon dioxide-rich fluid and hydrate formation in mid-Okinawa trough backarc basin. Science 248: 1093–1096.
Schrenk MO, Kelley DS, Delaney JR, Baross JA . (2003). Incidence and diversity of microorganisms within the walls of an active deep-sea sulfide chimney. Appl Environ Microbiol 69: 3580–3592.
Summit M, Baross JA . (1998). Thermophilic subseafloor microorganisms from the 1996 North Gorda Ridge eruption. Deep Sea Res Part II Top Stud Oceanogr 45: 2751–2766.
Summit M, Peacock A, Ringelberg D, White DC, Baross JA . (2000). Phospholipid fatty acid-derived microbial biomass and community dynamics in hot, hydrothermally influenced sediments from Middle Valley, Juan De Fuca Ridge. Proc Ocean Drill Prog Sci Results 169: 1–19.
Takai K, Gamo T, Tsunogai U, Nakayama N, Hirayama H, Nealson KH et al. (2004). Geochemical and microbiological evidence for a hydrogen-based, hyperthermophilic subsurface lithoautotrophic microbial ecosystem (HyperSLiME) beneath an active deep-sea hydrothermal field. Extremophiles 8: 269–282.
Takai K, Komatsu T, Inagaki F, Horikoshi K . (2001). Distribution of archaea in a black smoker chimney structure. Appl Environ Microbiol 67: 3618–3629.
Takai K, Mottl MJ, Nielsen SH,, The Expedition 331 Scientists. (2011) Proceedings of IODP, 331. Integrated Ocean Drilling Program Management International, Inc.: Washington, DC, USA.
Takai K, Nakagawa S, Reysenbach A, Hoek J . (2006) Microbial ecology of mid-ocean ridges and back-arc basins. In: Christie D et al. (eds). Geophysical Monograph Series. American Geophysical Union: Wasshington, DC, USA pp 185–213.
Takai K, Nakamura K, LaRowe D, Amend JP . (2014) Life at Subseafloor Extremes. In: Stein R, et al. (eds). Earth and Life Processes Discovered from Subseafloor Environments—A Decade of Science Achieved by the Integrated Ocean Drilling Program (IODP). Elsevier: Amsterdam, pp 149–174.
Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, Miyazaki J et al. (2008). Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc Natl Acad Sci USA 105: 10949–10954.
Takai K, Nakamura K . (2011). Archaeal diversity and community development in deep-sea hydrothermal vents. Curr Opin Microbiol 14: 282–291.
Tasumi E, Yanagawa K, Miyazaki J, Takai K . (2015) In vitro high-pressure incubation and activity measurement of deep-sea methanogenic archaea. In: McGenity TJ, et al. (eds) Hydrocarbon and Lipid Microbiology Protocols. Springer: Berlin Heidelberg, pp 1–14.
Teske A, Callaghan AV, LaRowe DE . (2014). Biosphere frontiers of subsurface life in the sedimented hydrothermal system of Guaymas Basin. Front Microbiol 5: 362.
Teske A, De Beer D, McKay L, Tivey MK, Biddle JF, Hoer D et al. (2016). The Guaymas Basin hiking guide to hydrothermal mounds, chimneys and microbial mats: complex seafloor expressions of subsurface hydrothermal circulation. Front Microbiol 7: 75.
Teske A, Hinrichs K-U, Edgcomb V, de Vera Gomez A, Kysela D, Sylva SP et al. (2002). Microbial diversity of hydrothermal sediments in the Guaymas Basin: evidence for anaerobic methanotrophic communities. Appl Environ Microbiol 68: 1994–2007.
Toki T, Tsunogai U, Gamo T, Kuramoto S, Ashi J . (2004). Detection of low-chloride fluids beneath a cold seep field on the Nankai accretionary wedge off Kumano, south of Japan. Earth Planet Sci Lett 228: 37–47.
Umezawa T, Aoki S, Nakazawa T, Morimoto S . (2009). A high-precision measurement system for carbon and hydrogen isotopic ratios of atmospheric methane and its application to air samples collected in the western Pacific region. J Meteorol Soc Jpn 87: 365–379.
Von Damm KL, Bischoff JL . (1987). Chemistry of hydrothermal solutions from the southern Juan de Fuca Ridge. J Geophys Res Solid Earth 92: 11334–11346.
Wankel SD, Adams MM, Johnston DT, Hansel CM, Joye SB, Girguis PR . (2012). Anaerobic methane oxidation in metalliferous hydrothermal sediments: influence on carbon flux and decoupling from sulfate reduction. Environ Microbiol 14: 2726–2740.
Yanagawa K, Breuker A, Schippers A, Nishizawa M, Ijiri A, Hirai M et al. (2014). Microbial community stratification controlled by the subseafloor fluid flow and geothermal gradient at the Iheya North Hydrothermal Field in the mid-Okinawa Trough (Integrated Ocean Drilling Program Expedition 331). Appl Environ Microbiol 80: 6126–6135.
Yanagawa K, Morono Y, de Beer D, Haeckel M, Sunamura M, Futagami T et al. (2013a). Metabolically active microbial communities in marine sediment under high-CO2 and low-pH extremes. ISME J 7: 555–567.
Yanagawa K, Nunoura T, McAllister S, Hirai M, Breuker A, Brandt L et al. (2013b). The first microbiological contamination assessment by deep-sea drilling and coring by the D/V Chikyu at the Iheya North hydrothermal field in the Mid-Okinawa Trough (IODP Expedition 331). Front Microbiol 4: 327.
Yanagawa K, Sunamura M, Lever MA, Morono Y, Hiruta A, Ishizaki O et al. (2011). Niche separation of methanotrophic archaea (ANME-1 and -2) in methane-seep sediments of the eastern Japan Sea offshore Joetsu. Geomicrobiol J 28: 118–129.
Yoshida-Takashima Y, Nunoura T, Kazama H, Noguchi T, Inoue K, Akashi H et al. (2012). Spatial distribution of viruses associated with planktonic and attached microbial communities in hydrothermal environments. Appl Environ Microbiol 78: 1311–1320.
Acknowledgements
We are deeply grateful to the Integrated Ocean Drilling Program, the captains, offshore installation managers, operation superintendents, all the crew, the science party, expedition staff and technicians on Chikyu from Mantle Quest Japan, CDEX and JAMSTEC, who assisted us in drilling, coring, sampling, installation and measurements during IODP Expedition 331. We also thank the proponents of and all involved in the preparation of IODP Proposal 601. We are grateful to Uta Konno for technical support in the stable isotope analysis. Jay Quade provided constructive comments and helpful suggestions. This work was supported in part by the Grant-in-Aid for Scientific Research: Project TAIGA (New Scientific Research on Innovative Areas, 20109003), a Grant-in-Aid for Young Scientists (15H05335) to KY and by the German Research Foundation (DFG) priority program 527 IODP/ODP Grant SCHI 535/12 to AS.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on The ISME Journal website
Supplementary information
Rights and permissions
About this article

Cite this article
Yanagawa, K., Ijiri, A., Breuker, A. et al. Defining boundaries for the distribution of microbial communities beneath the sediment-buried, hydrothermally active seafloor. ISME J 11, 529–542 (2017). https://doi.org/10.1038/ismej.2016.119
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2016.119
This article is cited by
-
Deep-sea bottom-water environment change caused by sediment resuspension on the continental slope off Sanriku, Japan, before and after the 2011 Tohoku Earthquake
Progress in Earth and Planetary Science (2022)
-
Interactions between temperature and energy supply drive microbial communities in hydrothermal sediment
Communications Biology (2021)
