
Abstract
The nicotinamide adenine dinucleotide phosphate (NADPH) oxidase produces superoxide, thus regulating redox state in the vessel wall. Three single-nucleotide polymorphisms (SNPs; −930A/G, A640G and C242T) of the p22phox subunit have been associated with hypertension; however, their role in peripheral and central pressures in normotensive individuals has not been addressed. A total of 210 healthy, normotensive individuals were studied. Genotypes for the −930A/G, A640G and C242T polymorphisms were determined by polymerase chain reaction. Peripheral pressures were measured by mercury sphygmomanometer and aortic pressures by a validated device using applanation tonometry. Both peripheral and central pressures differed across −930A/G genotypes. G allele carriers showed higher levels of peripheral systolic blood pressure (PSBP; AA: 113±12, GG/AG: 119±12 mm Hg; P<0.01) and peripheral diastolic blood pressure (AA: 70±9, GG/AG: 73±10 mm Hg; P<0.05). Regarding central pressures, AA homozygotes had lower central systolic blood pressure (CSBP; AA: 103±12, GG/AG: 108±12 mm Hg; P<0.01) and central diastolic blood pressure (AA: 71±9, GG/AG: 74±10 mm Hg; P<0.05). In multiple linear regression analysis, presence of the G allele (AG or GG) independently predicted CSBP. Blood pressure levels did not differ across A640G and C242T genotypes. The −930A/G polymorphism of p22phox is a determinant of peripheral and central pressures in normotensive individuals. The G allele is associated with higher blood pressure in the brachial artery, as well as in the aorta. These findings further elucidate the role of this polymorphism in the regulation of blood pressure. In contrast, the A640G and C242T SNPs do not influence peripheral and central pressures in normotensives.
Similar content being viewed by others


Introduction
Excess production of reactive oxygen species (ROS) in the vascular milieu has been linked to the development and progression of hypertension.1 The nicotinamide adenine dinucleotide phosphate (NADPH) oxidase is a membrane-bound enzyme responsible for the production of ROS. A number of studies have established its role as a major source of vascular superoxide anion (•O2−) in the hypertensive setting.2, 3
The role of this enzyme has been the subject of extensive genetic research, and a number of single-nucleotide polymorphisms (SNPs) of the genes that encode its subunits have been reported to confer increased propensity for elevated levels of blood pressure.4, 5 The CYBA gene encodes the essential p22phox subunit of NADPH oxidase; substitution of adenosine (A) for guanosine (G) in position −930 or 640, as well as cytidine (C) for thymidine (T) in position 242 have been reported to be more frequent among hypertensives, suggesting a possible role for these SNPs in the development of essential hypertension.3, 6
However, a putative contribution of these polymorphisms to the regulation of blood pressure in a non-hypertensive setting has not yet been addressed. Furthermore, correlations with central blood pressures have not been investigated. Central aortic pressures are physiologically more relevant than peripheral pressures, accurately reflecting the afterload of the left ventricle. Furthermore, they predict cardiovascular risk independently of brachial pressures.7 Antihypertensive medications change peripheral and central pressures in a disparate manner and this may explain superiority of specific classes of agents in outcome trials.8, 9 To this end, we sought to investigate a possible contribution of the −930A/G, A640G and C242T SNPs of the p22phox subunit of NADPH oxidase to arterial blood pressure levels in a population of healthy, normotensive individuals. The effect on both peripheral (brachial) and central (aortic) pressures was studied in addition to the effect on arterial wave reflections.
Methods
Study population
The study population consisted of 210 Caucasian subjects of Greek origin, who were randomly selected from the employees of two large industries producing food and cosmetics. They were from various socioeconomic categories, performing office and manual work.
Participants with coronary artery disease, cerebrovascular disease, peripheral artery disease, systemic inflammatory disease and on medication (anti-inflammatory agents, including aspirin and corticosteroids, anticoagulants) were excluded from the study. Exclusion criteria also included hypertension (values of blood pressure, as measured on two different occasions, >140/90 mm Hg, or use of antihypertensive drugs), diabetes mellitus (fasting glucose >126 mg per 100 ml, or use of antidiabetic agents) and dyslipidemia (history or use of lipid-lowering agents).
Study design
The study was conducted in the morning in a quiet, temperature-controlled room at 23 °C. Subjects underwent a standardized medical history and medical examination, anthropometric measurements, laboratory tests and a 12-lead electrocardiogram and examination after abstaining from caffeine, ethanol and flavonoid-containing beverages for at least 12 h before the study. Blood pressure was measured twice on the right arm with a mercury sphygmomanometer and the average of two blood pressure values spaced by 1 min was used in the analyses. The first and fifth phase of the Korotkoff sounds were used to identify systolic blood pressure (SBP) and diastolic blood pressure (DBP), respectively. Pulse pressure was calculated as SBP minus DBP. After a 20-min rest period in the supine position, baseline measurements of arterial function indices were taken, followed by blood sampling for full blood count, glucose, creatinine and lipoprotein profile. Levels of oxidized low-density lipoprotein (oxLDL) were measured in a subgroup of our population (n=105) using a competitive enzyme-linked immunoassay (Mercodia AB, Uppsala, Sweden). Weight and height were measured and body mass index was calculated. Estimated glomerular filtration rate was calculated using the Cockroft–Gault equation. All participants gave their informed consent to participate in the study, which was approved by the local ethics committee.
Measurement of aortic pressures and wave reflection indices
Central pressures were calculated using a validated, commercially available system (SphygmoCor; AtCor Medical, Sydney, Australia), which uses the principle of applanation tonometry and appropriate acquisition and analysis software for non-invasive recording and analysis of the arterial pulse, as previously described.7, 10, 11 The radial pulse wave was transformed into the central pulse wave of the aorta through pulse wave analysis and use of a validated transfer function. Mean blood pressure was calculated as the integral of the aortic pressure waveform.
The augmentation index (AIx) of the central (aortic) pressure waveform was measured as a composite measure of the magnitude of wave reflections and arterial stiffness, which affects the timing of wave reflections.12 Augmented pressure is the pressure added to the incident wave by the returning reflected wave, and represents the increased afterload that the left ventricle must cope with. AIx is defined as augmented pressure divided by central pulse pressure and is expressed as a percentage.7, 10, 11 Larger values of AIx indicate increased wave reflections from the periphery and/or earlier return of the reflected wave as a result of increased pulse wave velocity (because of increased arterial stiffness), and vice versa. AIx was averaged from 10 to 12 successive waves and was corrected for a steady heart rate of 75 beats min–1.13
Genotyping
Peripheral blood was drawn, collected in EDTA and stored at −20 °C. DNA was extracted using a commercially available kit (QIamp DNA Blood Midi Kit; Qiagen, Valencia, CA, USA). The −930A/G, A640G and C242T CYBA gene polymorphisms were analyzed in all subjects using restriction fragment length polymorphism as previously described.6, 14 Genotypes were determined according to the pattern of the bands on electrophoresis by two independent investigators.
Statistical analysis
The Hardy–Weinberg equilibrium was tested using χ2-test. Statistical normality of the data was checked using the Kolmogorov–Smirnov test. The distributions of high-density lipoprotein, triglyceride and oxLDL values were skewed; they were log10 transformed and became normally distributed. Continuous variables are presented as mean±s.d. Categorical variables are reported as frequencies and group percentages. A test for linearity was used to evaluate the trend of the investigated variables. Differences across study groups were examined using Student's t-test or one-way between-group analysis of variance followed by Bonferonni correction for continuous variables; the χ2-test for independence was used for comparison of categorical variables. Seven multiple linear regression analyses with stepwise selection were conducted to determine the variables independently associated with peripheral, central and mean blood pressures. Peripheral systolic and diastolic blood pressure, peripheral pulse pressure, central systolic and diastolic blood pressure, central pulse pressure and mean blood pressure were the dependent variables of each model, respectively. Independent variables of the models included presence of the G allele (AG or GG genotypes) for the −930A/G SNP, age, sex, smoking, body mass index, total cholesterol and estimated glomerular filtration rate. All statistical analyses were performed using SPSS 13.0 (SPSS, Chicago, IL, USA). A two-tailed P-value of <0.05 was regarded as significant.
Sample size calculation was based on the hypothesis that a difference of 10 mm Hg in PSBP would be detected across genotypes. Previous studies of our group have shown an s.d. for PSBP of 13 mm Hg. On the basis of these assumptions, 32 subjects per genotype group (96 in total) were required to provide the study with 80% power at the 5% level of significance. We recruited additional subjects (n=210) for confidence.
Results
The characteristics of the study population are shown in Table 1. Distribution of alleles was consistent with the Hardy–Weinberg equilibrium for all SNPs (χ2=2.74, P=0.10; χ2=0.39, P=0.53; and χ2=0.06, P=0.80 for −930A/G, A640G and C242T, respectively).
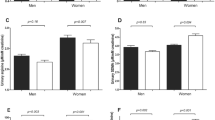
For −930A/G, the G allele frequency was 0.50 and the prevalence of AA, AG and GG genotypes was 27.6, 44.3 and 28.1%, respectively. Both PSBP and CSBP differed between the three genotypes, with AA subjects having lower pressures than GG and AG subjects. Presence of the G allele (GG/AG genotypes) resulted in higher peripheral (PSBP, peripheral diastolic blood pressure and peripheral pulse pressure) and central pressures (CSBP, central diastolic blood pressure and central pulse pressure) compared with the AA genotype. Mean blood pressure was similarly lower in AA homozygotes (Figure 1). No differences were detected regarding AIx; oxLDL levels did not differ either. In multivariate linear regression analyses, presence of the G allele of the −930A/G polymorphism was an independent predictor of CSBP levels (Table 2).

Blood pressure levels for −930A/G genotypes. *P<0.05; **P<0.01. SBP, systolic blood pressure; DBP, diastolic blood pressure; PP, pulse pressure; MBP, mean blood pressure.
For the A640G polymorphism, the G allele frequency was 0.47 and the prevalence of AA, AG and GG genotypes was 29.5, 47.6 and 22.9%, respectively. For the C242T polymorphism, the T allele frequency was 0.42 and the prevalence of CC, CT and TT genotypes was 33.3, 49.5 and 17.2%, respectively. Neither peripheral nor central blood pressure levels differ across genotypes of the two SNPs. Similarly, oxLDL levels were not different between groups.
Discussion
This is the first study, to the best of our knowledge, to explore the effect of the −930A/G, A640G and C242T polymorphisms of the CYBA gene on a healthy, normotensive population. In addition, the effect of these genetic loci on central pressures had not been addressed hitherto in any population, neither hypertensive nor normotensive. We have shown that carriage of the G allele of the −930A/G SNP is linked with higher, yet within the normal range, levels of both peripheral as well as central pressures.
NADPH activation may participate in the regulation of blood pressure through different mechanisms mediated by ROS. Indeed, upregulation of superoxide (•O2−) production leads to inactivation of nitric oxide. Moreover, peroxynitrite (ONOO−) and hydrogen peroxide (H2O2) production pave the way for media hypertrophy. NADPH oxidase, the key enzyme responsible for ROS production in arteries, consists of six subunits.15, 16, 17 The p22phox subunit is essential for the formation of a functional NADPH oxidase complex in all tissues. The CYBA gene, which encodes this subunit, has been shown to have several genetic polymorphic loci. Genetic variation has been linked to presence of hypertension,3 cerebrovascular disease18 and atherosclerosis.19 SNPs at positions −930, 640 and 242 of the gene modulate superoxide production and are linked to presence of hypertension as well as increased coronary artery disease risk among hypertensives.3, 4, 20 The −930A/G SNP is located at the promoter region of the gene, and regulates transcriptional activation. Hypertensive GG homozygotes show enhanced transcription of the CYBA gene, increased production of the p22phox subunit, upregulation of NADPH oxidase activity and, ultimately, increased ROS production.21 The A640G, located at the 3′-untranslated region of the gene, influences p22phox synthesis,22 whereas C242T modifies the binding of heme due to a replacement of histidine by tyrosine at amino acid position 72.19
Our findings regarding central pressures are particularly interesting. Aortic pressures are pathophysiologically more relevant than peripheral pressures, inasmuch as they are the pressures that the left ventricle must cope against.7 Prospective studies have established their role as markers and predictors of disease independently of brachial pressures. Moreover, corroborative data point to the differential effects of antihypertensive drugs at the aortic level despite similar effects at the periphery, and this may explain superiority in outcome of certain class of agents.8, 9, 16 Given the fact that the blood pressure-lowering agents that improve central hemodynamics also reduce oxidative load, we incorporated central blood pressures and wave reflection measurements in the design of our study. Indeed, the −930A/G SNP influenced aortic blood pressure levels, and GG/AG genotype independently predicted CSBP levels after adjusting for multiple confounders.
Interestingly, the role of the −930A/G polymorphism seems to extend beyond the regulation of blood pressure. It has been recently linked to propensity for insulin resistance,23 as well as increased carotid intima-media thickness in young smokers.24 Moreover, advanced age increases expression of p22phox, with a concomitant increase in superoxide anion production from the endothelium and the adventitia in a rodent model.25 Taken together, these results raise the possibility that carriers of the G allele may develop a greater risk of cardiovascular mishap on account of exposure to multiple vascular injurious stimuli.
Scant experimental data exist with regard to the role of the p22phox polymorphisms in normotensives. San José et al.21 have addressed the role of −930A/G in blood pressures both within and above the normal range. Although in hypertensive patients GG homozygotes produced excess superoxide, as a consequence of p22phox upregulation, protein levels and NADPH activity did not differ across the three genotypes in the normotensive group. This is in keeping with our finding that oxLDL levels did not differ among genotypes of the −930A/G, as well as for A640G and C242T. These findings when interpreted together with our results underscore the need for further research, incorporating •O2− measurements, in the subject.
Regarding dominance, the exact significance of G and A alleles for the −930A/G SNP, however, await clarification, because of the fact that non-uniform results are reported in various populations with different risk factors. In our study, presence of the G allele (GG/AG genotypes) conferred higher blood pressures among normotensives. In the setting of hypertension, only GG genotypes showed higher NADPH activation,21 whereas in young non-smokers it was the AA genotype that was associated with increased carotid intima-media thickness.24
The advent of NADPH oxidase inhibitors breaks new ground pertaining to potential clinical applications. Apocynin26, gp91ds-tat27 and protein kinase C inhibitors28 have been shown to decrease blood pressure; nevertheless, they have not been implemented in clinical practice.15 Dihydropyridines29 and blockade of the renin-angiotensin-aldosterone axis by means of angiotensin-converting inhibitors30 or angiotensin receptor blockers31 have been shown to effectively inhibit NADPH activation and concomitant production of ROS. The reduction of superoxide production through NADPH inhibition is ascribed as an additional pleiotropic effect of statins.32
A limitation of our study was that levels of superoxide anion, as an index of NADPH activity, were not measured; such analyses would require electron spin resonance spectrometry or a chemiluminescence assay. Similarly, mRNA or protein levels of p22phox were not determined. It should be emphasized that the population of our study consisted solely of Caucasians, and thus our results may not be directly extrapolated to different groups. Further research in large and diverse populations of various ethnic backgrounds and risk factor profiles is warranted to explore the effect of the CYBA gene SNPs and haplotypes on blood pressure levels.
In conclusion, we have shown that the −930A/G polymorphism at the promoter region of the CYBA gene that encodes the p22phox subunit of NADPH oxidase is independently correlated with brachial and aortic blood pressure levels in a normotensive population. Presence of the G allele confers higher blood pressure levels when compared with AA homozygotes. In contrast, the A640G and C242T SNPs do not influence peripheral and central blood pressures in normotensive individuals. These findings lend further support to the role of p22phox SNPs in blood pressure regulation.
References
Tain YL, Baylis C . Dissecting the causes of oxidative stress in an in vivo model of hypertension. Hypertension 2006; 48: 828–829.
Paravicini TM, Touyz RM . NADPH oxidases, reactive oxygen species, and hypertension: clinical implications and therapeutic possibilities. Diabetes Care 2008; 31 (Suppl 2): S170–S180.
Zalba G, San José G, Moreno MU, Fortuño A, Díez J . NADPH oxidase-mediated oxidative stress: genetic studies of the p22(phox) gene in hypertension. Antioxid Redox Signal 2005; 7: 1327–1336.
Moreno MU, José GS, Fortuño A, Beloqui O, Díez J, Zalba G . The C242T CYBA polymorphism of NADPH oxidase is associated with essential hypertension. J Hypertens 2006; 24: 1299–1306.
Calo LA, Davis PA, Pagnin E, Semplicini A, Pessina AC . NADPH oxidase, superoxide overproduction and nitric oxide bioavailability in essential hypertension. J Hypertens 2005; 23: 665–666.
Moreno MU, San José G, Orbe J, Páramo JA, Beloqui O, Díez J, Zalba G . Preliminary characterisation of the promoter of the human p22(phox) gene: identification of a new polymorphism associated with hypertension. FEBS Lett 2003; 542: 27–31.
Agabiti-Rosei E, Mancia G, O’Rourke MF, Roman MJ, Safar ME, Smulyan H, Wang J-G, Wilkinson IB, Williams B, Vlachopoulos C . Central blood pressure measurements and antihypertensive therapy: a consensus document. Hypertension 2007; 50: 154–160.
Vlachopoulos C, Hirata K, O’Rourke MF . Pressure-altering agents affect central aortic pressures more than is apparent from upper limb measurements in hypertensive patients: the role of arterial wave reflections. Hypertension 2001; 38: 1456–1460.
Williams B, Lacy PS, Thom SM, Cruickshank K, Stanton A, Collier D, Hughes AD, Thurston H, O’Rourke M, CAFE Investigators; Anglo-Scandinavian Cardiac Outcomes Trial Investigators; CAFE Steering Committee and Writing Committee. Differential impact of blood pressure-lowering drugs on central aortic pressure and clinical outcomes: principal results of the Conduit Artery Function Evaluation (CAFE) study. Circulation 2006; 113: 1213–1225.
Dima I, Vlachopoulos C, Alexopoulos N, Baou K, Vasiliadou C, Antoniades C, Aznaouridis K, Stefanadi E, Tousoulis D, Stefanadis C . Association of arterial stiffness with the angiotensin-converting enzyme gene polymorphism in healthy individuals. Am J Hypertens 2008; 21: 1354–1358.
Laurent S, Cockcroft J, Van Bortel L, Boutouyrie P, Giannattasio C, Hayoz D, Pannier B, Vlachopoulos C, Wilkinson I, Struijker-Boudier H, European Network for Non-Invasive Investigation of Large Arteries. Expert consensus document on arterial stiffness: methodological issues and clinical applications. Eur Heart J 2006; 27: 2588–2605.
Vlachopoulos C, Aznaouridis K, Stefanadis C . Clinical appraisal of arterial stiffness: the Argonauts in front of the Golden Fleece. Heart 2006; 92: 1544–1550.
Wilkinson IB, MacCallum H, Flint L, Cockcroft JR, Newby DE, Webb DJ . The influence of heart rate on augmentation index and central arterial pressure in humans. J Physiol 2000; 525 (Part 1): 263–270.
Inoue N, Kawashima S, Kanazawa K, Yamada S, Akita H, Yokoyama M . Polymorphism of the NADH/NADPH oxidase p22 phox gene in patients with coronary artery disease. Circulation 1998; 97: 135–137.
Williams HC, Griendling KK . NADPH oxidase inhibitors: new antihypertensive agents? J Cardiovasc Pharmacol 2007; 50: 9–16.
Lassegue B, Sorescu D, Szocs K, Yin Q, Akers M, Zhang Y, Grant SL, Lambeth JD, Griendling KK . Novel gp91(phox) homologues in vascular smooth muscle cells : nox1 mediates angiotensin II-induced superoxide formation and redox-sensitive signaling pathways. Circ Res 2001; 88: 888–894.
Touyz RM, Chen X, Tabet F, Yao G, He G, Quinn MT, Pagano PJ, Schiffrin EL . Expression of a functionally active gp91phox-containing neutrophil-type NAD(P)H oxidase in smooth muscle cells from human resistance arteries: regulation by angiotensin II. Circ Res 2002; 90: 1205–1213.
Genius J, Grau AJ, Lichy C . The C242T polymorphism of the NAD(P)H oxidase p22phox subunit is associated with an enhanced risk for cerebrovascular disease at a young age. Cerebrovasc Dis 2008; 26: 430–433.
Cahilly C, Ballantyne CM, Lim DS, Gotto A, Marian AJ . A variant of p22(phox), involved in generation of reactive oxygen species in the vessel wall, is associated with progression of coronary atherosclerosis. Circ Res 2000; 86: 391–395.
Gardemann A, Mages P, Katz N, Tillmanns H, Haberbosch W . The p22 phox A640G gene polymorphism but not the C242T gene variation is associated with coronary heart disease in younger individuals. Atherosclerosis 1999; 145: 315–323.
San Jose G, Moreno MU, Olivan S, Beloqui O, Fortuno A, Diez J, Zalba G . Functional effect of the p22phox −930A/G polymorphism on p22phox expression and NADPH oxidase activity in hypertension. Hypertension 2004; 44: 163–169.
Macias-Reyes A, Rodriguez-Esparragon F, Caballero-Hidalgo A, Hernandez-Trujillo Y, Medina A, Rodriguez-Perez JC . Insight into the role of CYBA A640G and C242T gene variants and coronary heart disease risk. A case-control study. Free Radic Res 2008; 42: 82–92.
Ochoa MC, Razquin C, Zalba G, Martinez-Gonzalez MA, Martinez JA, Marti A . G allele of the −930A>G polymorphism of the CYBA gene is associated with insulin resistance in obese subjects. J Physiol Biochem 2008; 64: 127–133.
Fan M, Raitakari OT, Kahonen M, Juonala M, Hutri-Kahonen N, Porsti I, Viikari J, Lehtimaki T . The association between cigarette smoking and carotid intima-media thickness is influenced by the −930A/G CYBA gene polymorphism: the Cardiovascular Risk in Young Finns Study. Am J Hypertens 2009; 22: 281–287.
Hamilton CA, Brosnan MJ, McIntyre M, Graham D, Dominiczak AF . Superoxide excess in hypertension and aging: a common cause of endothelial dysfunction. Hypertension 2001; 37 (2 Part 2): 529–534.
Baumer AT, Kruger CA, Falkenberg J, Freyhaus HT, Rosen R, Fink K, Rosenkranz S . The NAD(P)H oxidase inhibitor apocynin improves endothelial NO/superoxide balance and lowers effectively blood pressure in spontaneously hypertensive rats: comparison to calcium channel blockade. Clin Exp Hypertens 2007; 29: 287–299.
Rey FE, Cifuentes ME, Kiarash A, Quinn MT, Pagano PJ . Novel competitive inhibitor of NAD(P)H oxidase assembly attenuates vascular O(2)(−) and systolic blood pressure in mice. Circ Res 2001; 89: 408–414.
Beckman JA, Goldfine AB, Gordon MB, Garrett LA, Creager MA . Inhibition of protein kinase Cbeta prevents impaired endothelium-dependent vasodilation caused by hyperglycemia in humans. Circ Res 2002; 90: 107–111.
Rosenkranz AC, Lob H, Breitenbach T, Berkels R, Roesen R . Endothelial antioxidant actions of dihydropyridines and angiotensin converting enzyme inhibitors. Eur J Pharmacol 2006; 529: 55–62.
van der Giet M, Erinola M, Zidek W, Tepel M . Captopril and quinapril reduce reactive oxygen species. Eur J Clin Invest 2002; 32: 732–737.
Fortuno A, Bidegain J, Robador PA, Hermida J, Lopez-Sagaseta J, Beloqui O, Diez J, Zalba G . Losartan metabolite EXP3179 blocks NADPH oxidase-mediated superoxide production by inhibiting protein kinase C: potential clinical implications in hypertension. Hypertension 2009; 54: 744–750.
Whaley-Connell A, Habibi J, Nistala R, Cooper SA, Karuparthi PR, Hayden MR, Rehmer N, DeMarco VG, Andresen BT, Wei Y, Ferrario C, Sowers JR . Attenuation of NADPH oxidase activation and glomerular filtration barrier remodeling with statin treatment. Hypertension 2008; 51: 474–480.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article
Cite this article
Xaplanteris, P., Vlachopoulos, C., Baou, K. et al. The effect of p22phox −930A/G, A640G and C242T polymorphisms of NADPH oxidase on peripheral and central pressures in healthy, normotensive individuals. Hypertens Res 33, 814–818 (2010). https://doi.org/10.1038/hr.2010.78
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/hr.2010.78
Keywords
This article is cited by
-
The −930A>G polymorphism of the CYBA gene is associated with premature coronary artery disease. A case–control study and gene–risk factors interactions
Molecular Biology Reports (2014)
-
Endothelium-derived hyperpolarizing factor and hypertension
Hypertension Research (2013)
-
Association of NADPH oxidase p22phox gene C242T, A640G and −930A/G polymorphisms with primary knee osteoarthritis in the Greek population
Molecular Biology Reports (2013)
-
The NADPH Oxidase Family and its Inhibitors
Archivum Immunologiae et Therapiae Experimentalis (2012)
-
Association between the p22phox −930A/G polymorphism and blood pressure in normotensive subjects
Hypertension Research (2010)
