
Abstract
Purpose: Smith-Magenis syndrome (SMS), a probable contiguous gene syndrome due to an interstitial deletion of chromosome 17 band p11.2, is associated with a distinct and complex phenotype, including physical, developmental, and neurobehavioral features. The majority of SMS patients are deleted for a common ∼ 4 Mb interval that includes the gene SREBF1, a transmembrane transcription factor that regulates the low density lipoprotein (LDL) receptor and plays a crucial role in cholesterol homeostasis. A systematic study of fasting lipid profiles of patients with SMS was conducted to determine the frequency of cholesterol abnormalities.
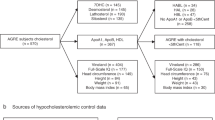
Methods: Fasting lipid profiles were examined in 49 children (27F/22M) between the ages of 0.6 years to 17.6 years (mean, 6.9 years) with a cytogenetically confirmed diagnosis of SMS. Observed values for serum total cholesterol (TC), triglycerides (TG), LDL cholesterol, and high density lipoprotein cholesterol were compared with published norms. The body mass index (BMI) was used as a measure of nutritional status.
Results: Mean TC was significantly higher than published NHANES III pediatric norms (P < 0.0008). Overall 28 of 49 (57%) SMS subjects had lipid values greater than the 95th percentile for age and gender for at least one or more of the following: TC, TG, and/or LDL. Only 16 SMS subjects (32%) were within normal limits for all three of these variables. BMI values showed minimal positive correlation to SMS lipid values; however, no consistent effect was found. Thus BMI values alone do not explain the marked trend in increased TC, TG, and/or LDL observed in the SMS group. Based on the American Academy of Pediatrics recommended lipid levels for children and adolescents, only one third of SMS subjects fall within normal range for TC and LDL; an additional one third each measure “borderline” or “high” for these values.
Conclusion: Hypercholesterolemia is common in SMS and may serve as a useful early clinical biochemical marker of the syndrome.
Similar content being viewed by others


Main
Smith-Magenis syndrome (SMS) is a complex multisystem, multiple congenital anomalies and mental retardation syndrome caused by an interstitial deletion of chromosome 17, band p11.2. Over 150 patients, ranging from 1 month to 72 years of age, have been identified worldwide from a diversity of ethnic groups. The deletion was first reported in 1982,1 and the phenotype more fully delineated in several large clinical series.2–10 Common features include craniofacial anomalies; infantile hypotonia with failure to thrive; otolaryngologic anomalies with speech delay, hoarse voice; short stature; brachydactyly; signs of peripheral neuropathy; and ophthalmologic, cardiovascular, and renal anomalies. Mental retardation is a feature, with most persons functioning in the moderate range of retardation.5 Also characteristic is the neurobehavioral phenotype, consisting of stereotypic, self-injurious and aggressive behaviors, and a significant sleep disturbance potentially due to an aberrant circadian rhythm.4,5,7–9,11–13
SMS is considered to be a contiguous gene syndrome as haploinsufficiency of multiple, functionally unrelated genes located in close proximity is thought to be responsible for the phenotype.4 Although the gene(s) responsible for SMS phenotype have yet to be determined, the majority of patients are deleted for a common interval spanning an estimated 4 to 5 Mb14,15; however, deletions can range from 2 to 9 Mb.16 Over 90% of SMS patients demonstrate a novel junction fragment by pulsed-field gel electrophoresis suggesting a common deletion.17
A gene involved in lipid metabolism, SREBF1 (sterol regulatory element binding protein 1), was mapped to 17p11.2.18 Subsequent physical mapping placed it within the SMS critical region.17,19SREBF1 is a transmembrane transcription factor involved in cholesterol homeostasis.20 In sterol-depleted cells, SREBF1 stimulates transcription of sterol regulated genes, including the low density lipoprotein (LDL) receptor and 3-hydroxy-3-methylglutaryl CoA reductase, the rate-limiting step in the cholesterol biosynthetic pathway. Through receptor-mediated endocytosis, the LDL receptor plays a crucial role in controlling the concentration of cholesterol in cells and plasma. Besides regulating the LDL receptor, these proteins coordinate the synthesis of two major components of membranes, fatty acids and cholesterol.21,22 The number of LDL receptors on the cell surface is under sterol feedback regulation.23 When intracellular concentration of cholesterol is high, LDL receptor gene transcription is repressed, with a concomitant rise in plasma LDL levels.
Because the SMS critical region contains SREBF1, patients with SMS are hemizygous for the gene encoding SREBP1. Preliminary evidence suggesting hypercholesterolemia is common in SMS was published in an abstract by Finucane et al.24 who found elevated total cholesterol in 11 of 13 (85%) SMS patients between the ages of 8 and 55 years. We conducted a systematic prospective study of lipid profiles in 49 SMS subjects < 19 years of age. Our results reveal a high frequency of hypercholesterolemia in SMS and suggest that haploinsufficiency for SREBF1 may affect cholesterol homeostasis in SMS.
MATERIALS AND METHODS
Study population
The study population consists of 49 children (27 female, 22 male) between the ages of 0.6 years to 17.6 years (mean, 6.9 years) with a cytogenetically confirmed diagnosis of SMS. All subjects underwent comprehensive interdisciplinary evaluation under institutional review board (IRB)–approved clinical protocols with required informed consent at one of two major research centers: the National Institutes of Health (NIH) or Texas Children's Hospital/Baylor College of Medicine (BCM). All had clinical exams consistent with SMS. The mean ages for the BCM and NIH groups were not statistically different (P = 0.1170), being 7.7 years and 5.7 years, respectively. De novo interstitial deletions of 17p11.2 were confirmed in all patients except two previously reported subjects: one male (BAB 1221) whose deletion of 17p11.2 was secondary to a de novo intrachromosomal rearrangement of chromosome 17, ins(17)(p11.2q11.2q21.3)25 and one female (BAB 484) with a larger deletion inherited from her mother, who was mosaic for del(17)(p11.2).26 Three patients (2 female, 1 male) were studied at both the NIH and BCM sites at different ages. To eliminate any statistical bias, only one set of cholesterol values was randomly selected and included in the analysis. None of the SMS subjects had any clinical signs of hypercholesterolemia, such as tendinous xanthomas or corneal arcus. Echocardiograms performed did not reveal evidence of ischemic cardiovascular disease. Although BCM methodology to measure serum triglyceride levels are not blanked for free glycerol, none of SMS subjects had evidence of diabetes, a major cause of elevated free glycerol, thereby minimizing the likelihood of falsely high triglyceride levels. Because hypothyroidism can cause elevations of total cholesterol and LDL cholesterol, thyroid function was also assessed by measuring thyroid-stimulating hormone (TSH) and free thyroxin.
Laboratory analysis of cholesterol
Blood was drawn for cholesterol profile after overnight fasting. Direct total cholesterol (TC), LDL cholesterol, high density lipoprotein (HDL) cholesterol, and triglycerides (TG) were measured in serum by standard methodologies in accordance with each center's protocol. Guidelines published by the Centers for Disease Control (CDC) has led to considerable standardization of methodologies used in lipid analysis. Texas Children's Hospital uses the VITROS test methodology manufactured by Ortho-Clinical Diagnostics. NIH uses the Sigma Cardiolipid reagents run on a COBAS FARA II and participates in the CDC Cholesterol Reference Method Laboratory Network (CRMLN) to ensure standardization to reference guidelines. Both NIH and BCM use methods that standardize against the same cholesterol reference (Abell-Kendall Reference Method).
Control population
A normal control population was not feasible for this study due to research constraints and consent requirements for testing in children. We elected to use published lipid reference levels as surrogates for a control population in this study. Despite considerable attention and agreement on the clinical guidelines for cholesterol testing and management in adults published by the National Cholesterol Education Program (NCEP) Expert Panel,27 normative lipid data for children remains limited. No single published reference range covers the ages of SMS study group (6 months–19 years). Therefore, observed SMS values were compared with published age and gender reference mean and percentile (5th, 50th, 95th) data derived from a few primary sources.
-
NHANES III: The third National Health and Nutrition Examination Survey (NHANES III) is a national survey conducted by the Centers for Disease Control (CDC) and Prevention's National Center for Health Statistics (NCHS) to produce national estimates on the nation's health and nutritional status.28 NHANES III includes survey data from 7,499 examinees collected from 1988 to 1994 and used to estimate mean and percentile distributions of serum TC, LDL, HDL, and TG in children and adolescents aged 4 to 19 years. Serum TC measurements in NHANES were standardized using the Abell-Kendall method, the same methodology used at NIH.
-
Clinical Guide to Lab Tests (3rd edition)29: This publication includes test methodology and reference range data for variety of general clinical tests, including lipid profiles of TC, LDL and HDL cholesterol, and TG. Reference lipid values for the 5th and 95th percentile for age and gender were derived from the Lipid Research Clinics (LRC) Prevalence Study.
-
Lipid Research Clinics Prevalence Study30: The LRC reference lipid data includes 5th, 50th, and 95th percentile values by age/gender.
Body mass index
Nutritional status and obesity among children and adolescents have been associated with adverse lipid levels. Because dietary information was not collected in a systematic manner for this study, the body mass index (BMI) value was used as a measure of SMS nutritional status.31 The BMI or Quetelet index, a measure widely used as a surrogate for obesity among children, is believed to account for 50% of the variability in body fat. Height and weight measurements on SMS study subjects taken at the time of lipid analysis were used to calculate BMI values by the formula that incorporates weight (W) and height (H) raised to a power: BMI = W/Ht2.31 Comparative statistical analysis by age and gender was conducted to examine the relationship, if any, between SMS BMI values and lipid profile results. BMI norms for subjects under 4 years of age required extrapolation from published curves32; all other norms were derived from published tables.33
Statistical analysis
Statistical analysis was systematically carried out to assess any differences between the mean lipid values for the BCM and NIH subject groups, before combining data from both groups. A Student t test was used for comparison of SMS lipid means to published age/gender reference means derived from NHANES III.28 Means were compared in the aggregate and by NHANES III age groups (4–5 years; 6–8 years; 9–11 years; and 12–19 years). SMS subjects < 4 years of age were compared with gender norms for the closest published reference age, e.g., 0 to 4 years SMS TC and HDL values were compared with normative mean values for 4 to 5 years NHANES III. LDL and TG normative means were not available for children under 12 years of age, restricting the statistical analysis to percentile observations. A second statistical test, the Arc Sine,34 was applied to examine the percentile distribution of lipid values, i.e., < 5th percentile; 5th to 95th percentile (normal range), and > 95th percentile. A standard Bonferroni correction for multiple testing was applied to all tests.35
RESULTS
Molecular analysis
Molecular and cytogenetic studies confirm hemizygosity for SREBF1 in all subjects, based on the presence of the SMS common deletion junction fragment, deletion for SREBF1 by FISH analysis, and/or prior published reports.15,25 Forty-five subjects have the common SMS deletion spanning ∼ 4 Mb. Three patients have larger deletions, two of which extend distally and include PMP22 (NIH 13; BAB 484)26 and the third male (BAB 536) previously reported by Juyal et al.15 One female (NIH 16) has a smaller deletion with more proximal breakpoints evidenced molecularly.
SMS fasting lipid profiles
Complete cholesterol profiles (TC, TG, LDL, and HDL) were available on 43 of 49 subjects; LDL and HDL were not available for 6 SMS subjects. There was no statistical difference between BCM and NIH means for TC (P = 0.93), LDL (P = 0.75), and HDL (P = 0.54). Mean TG values were somewhat higher (P = 0.02) in the BCM group (mean, 119.1) compared with the NIH group (mean, 85.5), but this finding was not significant after Bonferroni correction for multiple tests. Thus the BCM and NIH subjects were grouped together for all further analyses. A statistical comparison of observed mean lipid values in 49 SMS subjects compared with NHANES III population reference means is summarized in Table 1 and comparisons of SMS lipid values by age/gender reference percentiles in Table 2. A total of 18 statistical tests were performed in these two tables. Therefore, we have used 18 tests in our Bonferroni correction of significance levels for these tests.
Total cholesterol and triglycerides
SMS subjects had a mean TC value of 189.9 mg/dL (range, 116.0–267 mg/dL;N = 49), which was significantly higher than published population means (t = 3.34;P < 0.0008) (Table 1). Exploratory analysis by age group demonstrated a similar trend toward elevated TC at all age groups, except for SMS subjects between 6 and 11 years of age. The SMS group mean TG value was 104.7 mg/dL (range, 25.0–215 mg/dL). Statistical comparison of observed SMS TG values to published population means was restricted to SMS subjects aged 12 to 19 years. For this age group, the mean TG was 125.1 mg/dL and did not differ significantly from the NHANES III population means (t = 1.272;P < 0.20, not significant). Although statistical mean comparisons of TG below age 12 years were not possible, we noted that, at all younger groups, SMS mean TG SMS values were consistently higher than published 50th percentile norms.30 After Bonferroni correction, the comparison of mean TC values was still significant (P < 0.04).
LDL and HDL cholesterol
LDL and HDL cholesterol were measured on 43 of 49 SMS subjects (N = 43). SMS subjects had a mean LDL of 127.0 mg/dL (range, 64–194 mg/dL) (Table 1). Compared with the published NHANES III population mean for 12 to 19 years (mean, 95; SE 1.6), the SMS mean LDL, although increased for this age group (mean, 128.2; SE 13.2), was not quite significant (t = 1.876;P < 0.06) and was definitely not significant after Bonferroni correction (Table 1). Further statistical mean LDL comparisons were not possible for the younger age groups; however, SMS observed mean LDL values were consistently higher than the published LRC 50th percentile norms.30
The SMS mean HDL cholesterol was 42 mg/dL (range, 17–65 mg/dL). For all age groups, SMS subjects consistently had lower HDL values compared with NHANES III, but the difference was not statistically significant when analyzed by age group (Table 1). In the aggregate, SMS subjects' lower mean HDL was borderline significant compared with published population means (t = 1.923;P < 0.05) but not significant after correction for multiple tests.
Percentile distribution of SMS lipid levels
Observed SMS lipid values for TC, TG, LDL, and HDL were compared with published age/gender reference percentiles for the 5th and 95th percentiles (Table 2). Values were interpreted as low (below 5th percentile), normal (between the 5th and 95th percentile), or high (95th percentile and above). Although the majority of SMS subjects had normal TC (33 of 49; 67%), 14 of 49 (29%) of SMS subjects had high TC values compared with reference values. This finding represents a fivefold increase from expected numbers. Similarly, although normal TG levels were observed in 29 of 49 (59%), 39% (19 of 49) had high TG levels (> 95th percentile) for age and gender. Sixteen of 43 (37%) had high LDL cholesterol levels (95th percentile and above) for age. Although 86% of SMS subjects (37of 43) had HDL values in the normal range, 6 (14%) were low (below 5th percentile), almost three times the expected frequency.
Overall, 28 of 49 (57%) SMS subjects were greater than the 95th percentile for at least one or more of the following: TC, TG, and/or LDL. Sixteen SMS subjects (32%) were within normal limits for all three of these variables. Of the six subjects for whom LDL and HDL cholesterol values were not available, one had both elevated TG and TC; three had only elevated TG, and two had normal TG and TC.
Arc sine values
The Arc Sine (ts) test was used to examine the significance between the observed and expected number of subjects falling at or above the 95th percentile (Table 2). For a group size of 49, one would expect only 2.5 to fall below the 5th percentile, 2.5 above the 95th percentile, and 44 to be within the normal range (5th–95th percentile). The distribution of SMS subjects with lipid values at or above the 95th percentile deviated significantly from expected for TC (ts = 3.90, P < 0.00009), TG (ts = 5.16, P < 0.000001), and LDL (ts = 4.72;P < 0.000002), but not for HDL (ts = 1.72, P = 0.08). The first three tests remain significant after Bonferroni correction (P < 0.0016, 0.000018, and 0.000036, respectively).
Mean BMI analysis
The combined SMS mean BMI was 18.43 with a range of 14.08 to 31.67. There was no statistically significant difference found between the BCM and NIH groups (P = 0.36, not significant). The mean BMI for SMS females was 17.7 (SD, 3.65) and males (mean, 19.32; SD, 3.56). Although the BMI values for SMS females did not differ significantly from published norms (P = 0.11, not significant), SMS males were slightly higher (P < 0.001). Twelve SMS subjects had BMI values > 95th percentile for age/gender, which was significantly higher than expected by arc sine analysis (t = 3.36, P < 0.0008). BMI values were within the normal range (5th–95th percentile), for 33 of 49 (67%) SMS subjects. Four SMS subjects had BMI values < 5th percentile for age/gender. Among those with normal BMIs, 16 (48.5%) had TC, TG, and/or LDL values within the normal range; the remaining 17 were high for one or more of these. Only 9 of the 28 SMS subjects with elevated TC, LDL, and/or TG had high BMIs.
Correlation analysis of BMI and SMS lipid profile
To determine whether there was any association between BMI and lipid levels, correlation analysis was conducted. Results showed a small positive correlation between BMI and TC (r = 0.334;P = 0.02), LDL (r = 0.301;P = 0.05), and TG (r = 0.254;P = 0.08, not significant). However, no consistent relationship was identified between SMS lipid profile results and BMI scores for the 49 SMS study subjects, as demonstrated graphically for TC in Figure 1; similar graphs were found for TG, LDL, and HDL (data not shown). Linear regression (Fig. 2) was performed to determine whether BMI was a significant predictor of lipid levels in SMS subjects. BMI was significant for both TC (regression coefficient = 3.789;P < 0.02) and LDL (regression coefficient = 2.845;P < 0.05), but not for TG (regression coefficient = 3.481;P = 0.08, not significant) or HDL (regression coefficient = − 0.03;P = 0.94, not significant).

Correlation of TC (TotCHOL) and BMI. Of note is the variability of TC values for normal and high BMI values. Similar scattergrams found for LDL and TG (data not shown).

Linear regression analysis to determine the degree to which BMI predicts lipid values. BMI was significant for both TC (TotCHOL) (regression coefficient = 3.789;P < 0.019) (A) and LDL (regression coefficient = 2.845;P < 0.049) (B). Regression plots (not shown) were not significant for TG (regression coefficient = 3.481;P = 0.079, not significant) or HDL (regression coefficient = −0.03;P = 0.939, not significant).
DISCUSSION
Smith-Magenis syndrome (SMS), a probable contiguous gene syndrome caused by an interstitial deletion of chromosome 17p11.2, is characterized by a distinct and clinically recognizable phenotypic pattern of physical, developmental, and neurobehavioral features. Although the gene(s) responsible for SMS have yet to be determined, the majority of patients are deleted for a common deletion interval, SMS critical region, within 17p11.2 rendering them hemizygous for the genes within this interval. Parental origin of the 17p deletion has not been documented to affect the phenotype, suggesting that imprinting does not play a role in the expression of the Smith-Magenis syndrome phenotype.4,15 Although there is no evidence to suggest parental imprinting of the SREBF1 gene, this has not been conclusively demonstrated. Phenotypic variation can potentially arise from (1) variation in deletion size, (2) unmasking of a recessive allele (mutation) in patients with a corresponding deletion, or (3) haploinsufficiency for dosage-sensitive genes.36
Herein we describe abnormalities in fasting serum lipid profiles in 49 SMS children under 19 years of age who are hemizygous for SREBF1. In the aggregate, SMS mean TC was significantly higher than population means (P < 0.0008); age group comparisons also remained significant for age 5 and under and ages 12 to 19 years. Despite the suggestion of this age effect, the lack of population means for LDL and TG younger age groups does not permit further analysis of this trend. In the absence of published pediatric LDL and TG means, observed SMS lipid values were compared with published percentile norms based on NHANES III37 and/or the LRC.30 In this final analysis, SMS subjects deviated significantly compared with published percentile norms for TC, LDL, and TG (Table 2). Twenty-eight of 49 (57%) SMS subjects have values significantly greater than the 95th percentile for at least one or more parameters (TC, TG, and/or LDL). Only 16 SMS subjects (32%) were within normal limits for all three of these variables. Furthermore, although the majority had normal HDL levels, 6 of 43 (14%) were less than the 5th percentile for age/sex. Overall, there was at least a fivefold increase in the number of SMS subjects with high TC, TG, and/or LDL compared with expected numbers. These findings expand on a smaller preliminary report by Finucane et al.24 and confirm that SMS patients have a greater than expected frequency of hypercholesterolemia and hypertriglyceridemia.
Variables known to influence lipid levels were also examined to identify any potential bias for the elevated lipid values seen in these SMS subjects. Thyroid function was assessed in all but one male SMS subject whose TC and TG values were within normal range (BCM 536). Among the other 48 SMS subjects, 41 had normal thyroid function (TSH and Free T4), and 7 had normal TSH but low T4 values. Of the seven with low T4 values, only one had high TC but normal TG and LDL values. As expected, some of the variability for high TC and LDL relates to high BMI values, but not all of it. BMI was only weakly statistically significant in our regression analysis of TC and LDL (Fig. 2 A,B) and there were large numbers of children with high TC and LDL values who had low or normal BMIs. This finding suggests that nutritional status is not a major factor influencing lipid values in our sample. Among the 33 SMS subjects with normal BMI values, 16 had normal lipid values for TC, TG, and LDL; 17 of 33 (51.5%) were elevated for one or more. Because parental or sibling lipid levels were not a part of the IRB-approved research protocols, we cannot comment on the degree to which familial factors affect SMS lipid levels.
Because hypercholesterolemia was seen in over half of our SMS subject population, we considered the relevance of this finding to normal pediatric practice. Based on the American Academy of Pediatrics37 recommended lipid levels for children and adolescents (Table 3), only approximately one third of SMS subjects fall in the acceptable/desirable range for TC and LDL; one third fall in the borderline range, and over one third in the high range. SMS is a rare disorder with the majority of cases often not diagnosed until 4 to 5 years of age, when the behavioral aspects and craniofacial appearance of the syndrome are more recognizable. Early identification of elevated lipid levels (hypercholesterolemia) in a child with developmental and speech delays suggests the need for further evaluation and consideration of SMS diagnosis.
SMS patients are hemizygous for SREBF1 and would be predicted to have only half the normal amount of the encoded protein. Thus SMS provides a unique model for study of the cholesterol biosynthetic pathway. The elevated cholesterol levels observed in SMS subjects may be due to several factors, including haploinsufficiency for SREBF1 and/or other unidentified genes in the SMS critical region, as well as other modifier genes. However, haploinsufficiency for SREBF1 would not by itself be expected to cause a decrease in HDL or increase in TG. Although the possibility of a direct correlation between haploinsufficiency for the SREBF1 and the elevated cholesterol levels observed in this group of SMS patients is very enticing, the nature of this relationship remains to be elucidated. Shimano et al.38 have generated heterozygous and homozygous mice with targeted disruptions of SREBF1. Heterozygous animals were phenotypically normal; however, 50% to 85% of null mice died in utero. Interestingly the surviving null animals appeared normal, yet had elevated levels of mRNAs for HMG-CoA synthetase and reductase, farnesyl diphosphate synthetase, and squalene synthetase.38 Preliminary studies on cultured fibroblasts from three SMS patients also showed elevated levels of mRNA for HMG-CoA reductase, farnesyl diphosphate synthetase, and squalene synthetase, all enzymes involved in cholesterol synthesis.39 Continued functional studies of SMS fibroblasts are planned to further characterize how SREBF1 hemizygosity contributes to the SMS phenotype. There are no reports in the literature to suggest that hypercholesterolemia leads to atherosclerosis in SMS. Longitudinal clinical studies of individuals with SMS are required to determine whether or not an abnormal lipid profile in childhood has future clinical relevance with regard to cardiovascular disease.
References
Smith ACM, McGavran L, Waldstein G, Robinson J . Deletion of the 17 short arm in two patients with facial clefts. Am J Hum Genet 1982; 34: 410A.
Smith ACM, McGavran L, Robinson J, Waldstein G, Macfarlane J, Zonona J, Reiss J, Lahr M, Allen L, Magenis E . Interstitial deletion of (17)(p11.2p11.2) in nine patients. Am J Med Genet 1986; 24: 393–414.
Stratton RF, Dobyns WB, Greenberg F, DeSana JB, Moore C, Fidone G, Runge GH, Feldman P, Sekhon GS, Pauli RM, others. Interstitial deletion of (17)(p11.2p11.2): report of six additional patients with a new chromosome deletion syndrome. Am J Med Genet 1986; 24: 421–432.
Greenberg F, Guzzetta V, Montes de Oca-Luna R, Magenis RE, Smith AC, Richter SF, Kondo I, Dobyns WB, Patel PI, Lupski JR . Molecular analysis of the Smith-Magenis syndrome: a possible contiguous-gene syndrome associated with del(17)(p11.). Am J Hum Genet 1991; 49: 1207–1218.
Greenberg F, Lewis RA, Potocki L, Glaze D, Parke J, Killian J, Murphy MA, Williamson D, Brown F, Dutton R, McCluggage C, Friedman E, Sulek M, Lupski JR . Multi-disciplinary clinical study of Smith-Magenis syndrome (deletion 17p11.). Am J Med Genet 1996; 62: 247–254.
Chen K-S, Potocki L, Lupski JR . The Smith-Magenis syndrome (del(17)(p11.2)): clinical review molecular advances. Ment Retard Dev Disabil Res Rev 1996; 49: 1207–1218.
Smith AC, Dykens E, Greenberg F . Sleep disturbance in Smith-Magenis syndrome (del 17 p11.). Am J Med Genet 1998; 81: 186–191.
Smith AC, Dykens E, Greenberg F . Behavioral phenotype of Smith-Magenis syndrome (del 17p11.). Am J Med Genet 1998; 81: 179–185.
Dykens EM, Smith AC . Distinctiveness correlates of maladaptive behaviour in children adolescents with Smith-Magenis syndrome. J Intellect Disabil Res 1998; 42: 481–489.
Allanson JE, Greenberg F, Smith AC . The face of Smith-Magenis syndrome: a subjective objective study. J Med Genet 1999; 36: 394–397.
Dykens EM, Finucane BM, Gayley C . Brief report: cognitive behavioral profiles in persons with Smith- Magenis syndrome. J Autism Dev Disord 1997; 27: 203–211.
Potocki L, Glaze D, Tan DX, Park SS, Kashork CD, Shaffer LG, Reiter RJ, Lupski JR . Circadian rhythm abnormalities of melatonin in Smith-Magenis syndrome. J Med Genet 2000; 37: 428–433.
De Leersnyder H, De Blois MC, Claustrat B, Romana S, Albrecht U, Von Kleist-Retzow JC, Delobel B, Viot G, Lyonnet S, Vekemans M, Munnich A . Inversion of the circadian rhythm of melatonin in the Smith-Magenis syndrome. J Pediatr 2001; 139: 111–116.
Guzzetta V, Franco B, Trask BJ, Zhang H, Saucedo-Cardenas O, Montes de Oca-Luna R, Greenberg F, Chinault AC, Lupski JR, Patel PI . Somatic cell hybrids, sequence-tagged sites, simple repeat polymorphisms, yeast artificial chromosomes for physical genetic mapping of proximal 17p. Genomics 1992; 13: 551–559.
Juyal RC, Figuera LE, Hauge X, Elsea SH, Lupski JR, Greenberg F, Baldini A, Patel PI . Molecular analyses of 17p11.2 deletions in 62 Smith-Magenis syndrome patients. Am J Hum Genet 1996; 58: 998–1007.
Trask BJ, Mefford H, van den Engh G, Massa HF, Juyal RC, Potocki L, Finucane B, Abuelo DN, Witt DR, Magenis E, Baldini A, Greenberg F, Lupski JR, Patel PI . Quantification by flow cytometry of chromosome-17 deletions in Smith-Magenis syndrome patients. Hum Genet 1996; 98: 710–718.
Chen K-S, Manian P, Koeuth T, Potocki L, Zhao Q, Chinault AC, Lee CC, Lupski JR . Homologous recombination of a flanking repeat gene cluster is a mechanism for a common contiguous gene deletion syndrome. Nat Genet 1997; 17: 154–163.
Hua X, Wu J, Goldstein JL, Brown MS, Hobbs HH . Structure of the human gene encoding sterol regulatory element binding protein-1 (SREBF1) localization of SREBF1 SREBF2 to chromosomes 17p11.2 22q13. Genomics 1995; 25: 667–673.
Elsea SH, Purandare SM, Adell RA, Juyal RC, Davis JG, Finucane B, Magenis RE, Patel PI . Definition of the critical interval for Smith-Magenis syndrome. Cytogenet Cell Genet 1997; 79: 276–281 [published erratum appears in Cytogenet Cell Genet 1998;81:67].
Wang X, Sato R, Brown MS, Hua X, Goldstein JL . SREBP-1, a membrane-bound transcription factor released by sterol-regulated proteolysis. Cell 1994; 77: 53–62.
Brown MS, Goldstein JL . A receptor-mediated pathway for cholesterol homeostasis. Science 1986; 232: 34–47.
Brown MS, Goldstein JL . The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997; 89: 331–340.
Goldstein JL, Brown MS . Regulation of the mevalonate pathway. Nature 1990; 343: 425–430.
Finucane B, Smith ACM, Elsea SH, Greenberg F, Patel PI . Hypercholesterolemia in patients with del(17)(p11.2) (Smith-Magenis syndrome). Am J Hum Genet 1996; 59( suppl): A350.
Park JP, Moeschler JB, Davies WS, Patel PI, Mohandas TK . Smith-Magenis syndrome resulting from a de novo direct insertion of proximal 17q into 17p11.2. Am J Med Genet 1998; 77: 23–27.
Zori RT, Lupski JR, Heju Z, Greenberg F, Killian JM, Gray BA, Driscoll DJ, Patel PI, Zackowski JL . Clinical, cytogenetic, molecular evidence for an infant with Smith-Magenis syndrome born from a mother having a mosaic 17p11.2p12 deletion. Am J Med Genet 1993; 47: 504–511.
Panel. Executive Summary of the Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). JAMA 2001; 285: 2486–2497.
Hickman TB, Briefel RR, Carroll MD, Rifkind BM, Cleeman JI, Maurer KR, Johnson CL . Distributions trends of serum lipid levels among United States children, adolescents ages 4–19 Years. Data from the Third National Health Nutrition Examination Survey. Prev Med 1998; 27: 879–890.
Tietz NW, Pruden EL, McPherson RA, Fuhrman SA . Clinical guide to laboratory tests, 3rd ed. Philadelphia: Saunders, 1995;
Lipid Research Clinics (LRC) prevalence study. Bethesda, MD: NIH; 1980. Publication 80-1527.
Freedman DS, Perry G . Body composition health status among children adolescents. Prev Med 2000; 31: S34–S53.
Behrman RE, Kliegman RM, Jenson HB . Nelson textbook of pediatrics, 16th ed. Philadelphia: Saunders, 2000.
Hammer LD, Kraemer HC, Wilson DM, Ritter PL, Dornbusch SM . Standardized percentile curves of body-mass index for children adolescents. Am J Dis Child 1991; 145: 259–263.
Sokal RR, Rohlf FJ . Biometry. The principle and practice of statistics in biological research. San Francisco: W.H. Freeman, 1969.
Lachin JM . Biostatistical methods. The assessment of relative risks. New York: John Wiley & Sons. 2000: 117–118.
Lupski JR . Genomic disorders: structural features of the genome can lead to DNA rearrangements human disease traits. Trend Genet 1998; 14: 417–422.
American Academy of Pediatrics. Cholesterol in childhood (RE9805). Pediatrics 1998; 101: 141–147.
Shimano H, Shimomura I, Hammer RE, Herz J, Goldstein JL, Brown MS, Horton JD . Elevated levels of SREBP-2 cholesterol synthesis in livers of mice homozygous for a targeted disruption of the SREBP-1 gene. J Clin Invest 1997; 100: 2115–2124.
Goker-Alpan O, Blancato J, Smith ACM, Gahl WA . Expression of cholesterol biosynthetic enzymes is upregulated in Smith-Magenis syndrome fibroblasts. Am J Hum Genet 1999; 65( suppl): A360.
Acknowledgements
This work was supported in part by grants from the National Institute of Child Health and Development (NICHD, K08HD01149 to L.P.), (HD 08010; HD 38534 to S.H.E.); the National Cancer Institute (P01CA75719 to J.R.L.); the NIH Clinical Center DIR Bench-to-Bedside Award (A.C.M.S.); the Baylor College of Medicine Mental Retardation Research Center (HD2406402); the Baylor Child Health Research Center (HD94021); and the Texas Children's Hospital General Clinical Research Center (M01RR00188). Ann C.M. Smith extends special thanks to the late Jeffrey M. Hoeg, MD, Section of Cell Biology, Molecular Disease Branch, NHLBI, NIH, and Douglas Wilkin, PhD, Medical Genetics Branch, NHGRI, NIH, whose support and suggestions were integral in the early design of this study. We also express our thanks to Alan Remaly, M.D., PhD, Clinical Pathology Department, W.G. Magnuson Clinical Center, NIH for technical input related to laboratory standardization for lipid analysis.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Smith, A., Gropman, A., Bailey-Wilson, J. et al. Hypercholesterolemia in children with Smith-Magenis syndrome: del (17)(p11.2p11.2). Genet Med 4, 118–125 (2002). https://doi.org/10.1097/00125817-200205000-00004
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1097/00125817-200205000-00004
Keywords
This article is cited by
-
Metabolic effects of the schizophrenia-associated 3q29 deletion
Translational Psychiatry (2022)
-
Behavioral disturbance and treatment strategies in Smith-Magenis syndrome
Orphanet Journal of Rare Diseases (2015)
-
Genetic Counseling, Activism and ‘Genotype‐First’ Diagnosis of Developmental Disorders
Journal of Genetic Counseling (2012)
-
Smith–Magenis syndrome
European Journal of Human Genetics (2008)
-
Mutations of RAI1, a PHD-containing protein, in nondeletion patients with Smith-Magenis syndrome
Human Genetics (2004)
