
Abstract
Lynch syndrome (LS) is an inherited cancer-predisposing disorder caused by germline mutations in the mismatch repair (MMR) genes. The high variability in individual cancer risk observed among LS patients suggests the existence of modifying factors. Identifying genetic modifiers of risk could help implement personalized surveillance programs based on predicted cancer risks. Here we evaluate the role of the telomerase (hTERT) rs2075786 SNP as a cancer-risk modifier in LS, studying 255 and 675 MMR gene mutation carriers from Spain and the Netherlands, respectively. The study of the Spanish sample revealed that the minor allele (A) confers increased cancer risk at an early age. The analysis of the Dutch sample confirmed the association of the A allele, especially in homozygosity, with increased cancer risk in mutation carriers under the age of 45 (relative riskLSca<45_AA=2.90; 95% confidence interval=1.02–8.26). Rs2075786 is associated with colorectal cancer (CRC) risk neither in the general population nor in non-Lynch CRC families. In silico studies predicted that the SNP causes the disruption of a transcription binding site for a retinoid receptor, retinoid X receptor alpha, probably causing early telomerase activation and therefore accelerated carcinogenesis. Notably, cancer-affected LS patients with the AA genotype have shorter telomeres than those with GG. In conclusion, MMR gene mutation carriers with hTERT rs2075786 are at high risk to develop a LS-related tumor at an early age. Cancer-preventive measures and stricter cancer surveillance at early ages might help prevent or early detect cancer in these mutation carriers.
Similar content being viewed by others


Introduction
Lynch syndrome (LS) (MIM 120435) is an autosomal-dominant inherited disorder caused by germline mutations in the DNA mismatch repair (MMR) genes. It is characterized by early-onset colorectal cancer (CRC) and an increased risk of upper gastrointestinal, urologic and gynecologic cancers.1 Several studies have estimated the cancer risks associated with mutations in the MMR genes, but these estimates vary substantially across the studies (20–80% lifetime risk).2, 3, 4, 5, 6, 7, 8, 9
There is a considerable variation in LS expression and no obvious gene-specific genotype/phenotype correlations have been demonstrated nor does there appear to be any relationship between the location of a mutation and type of disease. In addition to environmental factors, there is evidence suggesting the existence of genetic factors that somehow explain the variability in individual cancer risk.10 Identifying the genetic modifiers of risk can lead to an efficient stratification of mutation carriers based on their predicted risk, implying thus, a more appropriate clinical management based on personalized surveillance programs.
Most of the attempts to identify cancer-risk modifiers in LS have been based on the study of candidate genes and most of them have not been validated when tested in larger sample sizes.11 In contrast, two genetic variants previously identified in CRC genome-wide association studies (GWAS), rs16892766 and rs3802842, might modify cancer risk in LS families.12, 13, 14
Telomeres are located at the end of chromosomes and protect the chromosome ends from nucleolitic degradation, end-to-end fusions and irregular recombination, being thus critical for genome stability and integrity. Telomeres progressively shorten with each cell replication cycle. Telomere length anomaly appears to be one of the earliest and most prevalent genetic alterations in the multistep process of malignant transformation.15, 16 Telomerase catalyzes the de novo addition of telomeric repeat sequences onto chromosome ends and it is usually inactive in normal somatic cells, whereas its expression has been linked to increased susceptibility to tumorigenesis.17, 18, 19 Genetic variants located in genes involved in telomere maintenance, and in particular in hTERT (MIM 187270), the gene encoding the catalytic subunit of telomerase, have been associated with increased risk to cancer.20, 21, 22, 23, 24, 25, 26, 27 Among the better characterized cancer variants, the hTERT rs2075786 (c.2654+269G/A) SNP has been associated with lung cancer risk.28, 29 However, no studies have reported any association between variants located at the hTERT locus and the risk of CRC.
Here we evaluate hTERT rs2075786 as a modifier of cancer in LS patients in 255 MMR gene mutation carriers from Spain. Also, 675 mutation carriers from the Netherlands were analyzed. We also assessed its role as cancer-risk factor in a population-based case–control series and in non-LS familial cases.
Materials and methods
Subjects
Spanish sample
A total of 255 individuals from 101 LS families whose mutation carrier statuses were known were included in the study. They were assessed through the Hereditary Cancer Program of the Catalan Institute of Oncology from 1998 to 2010. All families were of Caucasian origin. Informed consent was obtained from all individuals. MMR mutation analysis was performed on genomic DNA extracted from peripheral blood lymphocytes. Large genomic alterations were studied using multiplex ligation-dependent probe amplification (SALSA MLPA Kits, MRC-Holland, Amsterdam, The Netherlands). Mutation screening was performed by direct sequencing after PCR amplification (primers and conditions available upon request). In all, 147 (57.6%) MMR gene mutation carriers had been diagnosed with a LS-related tumor, of whom 134 with CRC. The clinical characteristics of the subjects are detailed in Table 1.
Also, DNA from 277 sporadic CRC cases and 280 controls were obtained from a hospital-based case–control study (Bellvitge Colorectal Cancer Study). Cases were consecutive patients with a first diagnosis of colorectal adenocarcinoma attending the Bellvitge University Hospital in Barcelona. For the group of patients, the male/female ratio is 1.2 and the mean age is 66.7 (range: 23–91), and for the controls, 1.1 and 65.5 (range: 24–92), respectively. Details about the study population were published elsewhere.30
Dutch sample
A total of 675 MMR gene mutation carriers from 127 different families from the Dutch LS Registry were studied. Detailed description of the registry, individuals and DNA extraction method was published before.12 Information on the subjects has been recently updated and thus, follow-up data added. In all, 201 (29.8%) MMR gene mutation carriers had been diagnosed with a LS-related tumor, of whom 146 are with CRC. Table 1 summarizes the updated clinical characteristics of the subjects.
The Dutch case–control series consists of 324 CRC cases and 785 controls. Cases are probands assessed through a familial cancer clinic; therefore, they are suspected of genetic CRC susceptibility but with no germline mutations identified in the MMR genes. Regarding controls, 475 are healthy blood donors and 310 are healthy spouses of MMR gene mutation carriers.
MMR gene mutation carriers are derived from multiple-case families selected for genetic counseling and testing. In this situation, the disease status clearly affects the likelihood of testing, causing an overrepresentation of affected individuals. This observation is evident in the Spanish but not in the Dutch sample. The Dutch registry has been running longer (it was established in 1985) and follows large and complete LS families, whereas the Spanish series comes from a cancer hospital-based genetic counseling unit established in 1998, where a clear ascertainment bias towards the assessment of cancer-affected family members exists. Because of the differences in affected vs unaffected ratios, the studies were performed independently for the Spanish and Dutch samples.
Genotyping
The rs2075786 SNP was genotyped using the commercially available TaqMan assay C_15824034_10 (Applied Biosystems Inc., Foster City, CA, USA) following the manufacturer’s instructions. Reactions were performed in duplicate in the LightCycler 480 real-time PCR detection system (Roche Diagnostics GmbH, Mannheim, Germany). Genotype calling was performed automatically by the LightCycler 480 II software (Roche Diagnostics GmbH, Mannheim, Germany). Genotyping failed in 78 of the 2596 (3%) samples included in the study.
In silico analyses
Public genome browsers (Ensembl, NCBI and USCS genome browsers) were checked out to investigate the existence of transcription of noncoding RNAs from the region where rs2075786 is located. The presence of microRNA-binding sites was studied using the microRNA target-prediction algorithms provided by http://www.microrna.org. The strength of the pseudoexon splice sites was analyzed using three different prediction programs: www.fruitfly.org/seq_tools/splice.html, http://genes.mit.edu/burgelab/maxent/Xmaxentscan_scoreseq.html and http://ast.bioinfo.tau.ac.il/SpliceSiteFrame.html, as previously described.31 Putative binding sites for transcription factors were identified using PROMO 3.0, a web-based program that employs the TRANSFAC database version 8.3 to construct specific binding-site weight matrices for prediction of transcription factor-binding sites.32, 33
Relative telomere length assessment
Telomere length quantification was performed using a monochrome multiplex quantitative PCR method, as described by Cawthon with slight modifications (available upon request).34
Statistical analyses
Evaluation of the deviation of rs2075786 from the expected Hardy–Weinberg equilibrium was performed in controls from each population. The SNP was in Hardy–Weinberg equilibrium in controls from both populations (PSpanish=0.48; PDutch=0.93).
The association between genotypes and risk of CRC or LS-related tumors was evaluated using conditional logistic regression adjusting for familial clustering. The end points considered were: age at diagnosis of the LS-related cancer or CRC for affected individuals and age of last follow-up for unaffected individuals. Likelihood ratio tests were applied to assess statistical significance. In general, relative risks (RRs), 95% confidence intervals (CIs) and associated P-values were estimated for different models of inheritance and the best model was selected using the Akaike information criterion. The interaction of genotypes with age stratified at 45 years was assessed to identify anticipation effects. For case–control studies, odds ratios (ORs) and 95% CI were calculated using logistic regression adjusted for gender. Kruskal–Wallis rank-based test was used to compare telomere length among rs2075786 genotype groups, adjusting for age and gender. Statistical analyses were performed using R.
Results
Spanish study
No differences in rs2075786 genotype distributions were observed when comparing MMR gene mutation carriers affected with cancer, either LS-related or only CRC, and unaffected mutation carriers (Supplementary Table S1).
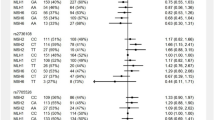
We further analyzed rs2075786 genotype distributions within two age intervals in the cancer group. For this aim, we established a cut off of 45 years of age based on the median age of all individuals included in the study. MMR gene mutation carriers diagnosed with cancer before 45 years of age were compared with those diagnosed after 45 years of age and those who were cancer-free after 45 years of age. Under this condition, the A allele conferred an increased risk of cancer in LS carriers under the age of 45 compared with those diagnosed with cancer or cancer-free after the age of 45 (Table 2).
Although borderline and not fulfilling the required standards for declaring an association, the results obtained in the Spanish sample led us to analyze another series (Dutch study) in a more comprehensive manner, thanks to the larger sample size and the availability of follow-up and polypectomy information.
Dutch study
A total of 675 MMR gene mutation carriers from the Dutch hereditary CRC registry were included in the analysis. As previously observed for the Spanish MMR gene mutation carriers, no effect was detected on the general risk of cancer between cancer-affected and unaffected individuals (Supplementary Table S2).
To study the effect on cancer risk related with age intervals, cancer-affected vs unaffected MMR gene mutation carriers were compared within the groups under 45 years and within the groups ≥45. For this purpose, the two age groups were created as: group age <45, MMR gene mutation carriers diagnosed with cancer under the age of 45 and unaffected MMR gene mutation carriers under age 45 and group age ≥45, cancer-affected and unaffected MMR gene mutation carriers >45. Because of the larger sample size and the fact that unaffected carriers had a similar or even higher average age than cancer-affected carriers (Table 1), this type of distribution in age groups could be performed for the Dutch MMR gene mutation carriers.
In this series, the AA genotype confers an increased risk of LS-related cancers in individuals <45 years of age (RR=2.76; 95% CI=1.02–7.50). When only CRC was considered, an RR of 2.46 (96% CI=0.78–7.82) was found for the AA homozygotes, but this observation was not statistically significant, probably because of the effect on the risk of all LS-related tumors. No association of rs2075786 with cancer risk was observed in the ≥45 groups (Table 3).
MMR gene mutation carriers are usually under strict clinical surveillance and undergo colonoscopies on a routine basis, implying that whenever detected, polyps are removed. As polypectomy affects subsequent CRC risk, we included this information in the analysis. Within the group age <45, all unaffected subjects with a polypectomy were excluded. Moreover, within the group age ≥45, all cancer-affected subjects with a polypectomy were performed under the age of 45 and all unaffected individuals with polypectomies were excluded from the analysis. A total of 100 individuals were removed. In this scenario, the increased cancer risk conferred by AA in the <45 group is maintained or even increased (RRLSca<45_AA=2.90; 95% CI=1.02–8.26) (Table 4). Slight random differences from the previous analysis dependent on the genotype of the cases excluded were observed.
No effect of gender on cancer risks was observed (data not shown). Regarding the MMR gene affected, the analysis was performed considering MLH1 and MSH2 separately (Supplementary Table S3). Although differences did not reach significance and larger series should be studied to draw definitive conclusions, the results suggest that the effect of rs2075786 on cancer risk occurs for both MLH1 and MSH2 mutation carriers under the age of 45 (RRMLH1<45_AA=3.86; 95% CI=0.70–21.33 and RRMSH2<45_AA=2.36; 95% CI=0.42–13.39).
Rs2075786 and CRC risk in the general population and in non-Lynch CRC families
To evaluate whether rs2075786 is associated with CRC risk in the general population and in non-Lynch familial CRC cases, we performed an association study in a case–control series from Spain and in non-Lynch CRC familial cases from the Netherlands. Rs2075786 was in Hardy–Weinberg equilibrium in both the Spanish (P=0.59) and Dutch (P=0.88) controls. No statistically significant association between rs2074786 and CRC was detected in any of the two situations, with ORs close to 1 (Supplementary Table S4).
Also, when separating by age groups (<45 and ≥45) in the non-Lynch familial CRC study (The Netherlands), no statistically significant differences were identified in either group (Supplementary Table S5). The age stratification could not be performed for the sporadic case–control series (Spain) because of the scarcity of sporadic CRC patients with <45 years of age present.
In silico studies to unravel rs2075786 functionality
We performed an in silico search of potential effects of the variant on gene splicing, transcription factor or microRNA binding, as well as of noncoding RNAs being transcribed from that genomic region. Public databases showed inexistence of noncoding RNAs, and microRNA target-prediction algorithms found no microRNA-binding sites when either allele (A or G) was present. Likewise, gene splicing was not affected. In silico search for putative binding sites of transcription factors revealed that when the rare allele (A) of rs2075786 is present, the elimination of a binding site for retinoid X receptor alpha (RXR-α) is predicted.
Rs2075786 and telomere length
To test the hypothesis that telomere length is compromised by the presence of the A allele of rs2075786, we measured the relative telomere length in blood DNA of the cancer-affected LS patients from Spain. Individuals with the AA genotype have shorter telomeres than individuals with the GG (P=0.011) and GA (P=0.058) genotypes (Figure 1). No association of relative telomere length with rs2075786 genotypes was found in sporadic CRC patients (Supplementary Figure S1).

Blood relative telomere length (RTL) in 146 cancer-affected LS patients according to rs2075786 genotype distribution (nGG=66, nAG=64 and nAA=16). The boxes represent the interquartile range of distributions (25th–75th percentile); the horizontal lines within the boxes, the medians; and the vertical lines, the 5th and 95th percentiles.
Discussion
There remains a considerable variability in LS disease expression that cannot be readily explained by genetic variance occurring solely within the MMR genes. Several studies have tried to determine whether there are any genetic modifying factors that could be associated with an increased likelihood of developing cancer. In this study, we show that the AA genotype of the variant rs2075786 of the telomerase gene (hTERT) is associated with an increased risk of LS-related cancers in MMR gene mutation carriers under 45 years of age, indicating anticipation in the age of onset of cancer. Moreover, when information about polypectomies, a therapeutic intervention that affects CRC risk and might lead to false negatives, was incorporated to the analyses, the results obtained improved, strengthening the observation (RR<45_AA=2.90; 95% CI=1.02–8.26).
The field of cancer-risk modifiers of LS is very active and has provided in the recent years controversial results. First, candidate genes that have some plausible biological role in the disease were evaluated. Except for rare exceptions, the initial findings identified in small-sized samples were not validated in larger series.11 More recently, some polymorphisms identified through GWAS of unselected CRC patients and controls have been shown to modify cancer risk in LS families. Wijnen et al12 first studied six GWAS CRC susceptibility SNPs in 675 MMR gene mutation carriers from 127 LS families. They found that two of them, at 8q23.3 and 11q23.1, were associated with CRC risk in mutation carriers, particularly in females.12 These results were partly confirmed in a cohort of Australian and Polish MMR mutation carriers, but were not replicated in a French cohort.13, 14
We selected the hTERT variant rs2075786 by somehow combining candidate gene and GWAS-based approaches. First, hTERT has an important role at very early stages of carcinogenesis, its expression has been linked to increased susceptibility to tumorigenesis, and rs2075786 has been associated with cancer risk.15, 16, 18, 19, 28, 29 And second, through GWAS, genetic variants located in genes involved in telomere maintenance, and in particular in hTERT, were also associated with increased risk to cancer.20, 21, 22, 23, 24, 25, 26, 27 In contrast to previous studies examining the role of modifier genes,11 our study includes two independent series of MMR gene mutation carriers, of 255 and 675 individuals, the latest being among the largest series used to study genetic modifiers of LS. In view of our results with rs2075786, it may be relevant to study the role of other hTERT polymorphisms as cancer-risk modifiers in LS, in particular those variants with functional implications and/or with strong levels of evidence of being associated with cancer risk.35
In order to unravel the functional effect of rs2075786, in silico analysis predicted that when the minor allele (A) is present, a binding site for RXR-α is eliminated. Already existent evidence indicates that retinoids, through retinoid receptors, inhibit telomerase activity and downregulate hTERT expression.36, 37 These observations suggest that in the presence of the rs2075786 minor allele A, where a binding site for retinoid receptors is absent, natural retinoids cannot efficiently restrain hTERT expression, causing accelerated tumor cell growth.
It is nevertheless intriguing that rs2075786 does not influence CRC risk in sporadic or in other non-Lynch familial cases. It is well known that the carcinogenic pathway of MMR-deficient tumors clearly differs from that involved in most sporadic and non-LS familial CRC. It has been observed that MMR-deficient cell lines and colon tumors show high mutation frequencies at telomere ends, leading to accelerated telomere shortening.38, 39, 40, 41 The effect of an increased telomere-shortening rate is likely to require early activation of telomerase in such tumors. As hypothesized above, the presence of the A allele of rs2075786 might imply even earlier activation of telomerase. Although the effect might be very subtle to have an impact in MMR-proficient cells, it could trigger important consequences in a MMR-deficient context, such as in LS tumors. In this line of research, it may be interesting to evaluate the role of rs2075786 as a risk allele for sporadic CRC with microsatellite instability.
A possible explanation for the rs2075786 acting as modifier of cancer risk at early ages but not later in life might be that the effect the variant has on telomere shortening (Figure 1) is not efficient at late ages where telomeres are already physiologically shortened due to the aging process.42 Nevertheless, further functional assays are required to provide experimental evidence about this matter.
When assessing small variations in cancer risk, the issue of personalizing follow-up immediately arises. A potential applicability of rs207586 genotyping in genetic counseling for clinical management of LS patients can be envisioned. Based on our results, young MMR gene mutation carriers with the rs2075786 AA genotype might benefit from an even more intensive clinical surveillance of CRC and other LS-related tumors than the one suggested by the standard LS surveillance protocols. Validation of our observation in additional series is warranted prior to translation to routine clinical practice. Nevertheless, both MMR gene mutation carriers and clinicians should be conscious of the higher risk that rs2075786 carriers have at early ages, observing or even increasing the screening frequency for colorectal, endometrial and other LS-related tumors. Also, lifestyle changes that affect environmental modifier risk factors might help reduce the risk of developing cancer. Likewise, the recent promising results of aspirin-based chemoprevention obtained in LS patients suggest that MMR gene mutation carriers with the rs2075786 AA genotype might benefit from this chemopreventive treatment starting at early age.42
In conclusion, we observe an association that might help identify MMR gene mutation carriers with higher risk to prematurely develop a LS-associated tumor. Rs2075786 adds up to the already known LS modifiers, increasing thus the opportunity to incorporate the information of modifier factors into clinical practice.
References
Lynch HT, Lynch PM, Lanspa SJ, Snyder CL, Lynch JF, Boland CR : Review of the Lynch syndrome: history, molecular genetics, screening, differential diagnosis, and medicolegal ramifications. Clin Genet 2009; 76: 1–18.
Dunlop MG, Farrington SM, Carothers AD et al: Cancer risk associated with germline DNA mismatch repair gene mutations. Hum Mol Genet 1997; 6: 105–110.
Aaltonen LA, Salovaara R, Kristo P et al: Incidence of hereditary nonpolyposis colorectal cancer and the feasibility of molecular screening for the disease. N Engl J Med 1998; 338: 1481–1487.
DeFrancisco J, Grady WM : Diagnosis and management of hereditary non-polyposis colon cancer. Gastrointest Endosc 2003; 58: 390–408.
Quehenberger F, Vasen HF, van Houwelingen HC : Risk of colorectal and endometrial cancer for carriers of mutations of the hMLH1 and hMSH2 gene: correction for ascertainment. J Med Genet 2005; 42: 491–496.
Jenkins MA, Baglietto L, Dowty JG et al: Cancer risks for mismatch repair gene mutation carriers: a population-based early onset case-family study. Clin Gastroenterol Hepatol 2006; 4: 489–498.
Alarcon F, Lasset C, Carayol J et al: Estimating cancer risk in HNPCC by the GRL method. Eur J Hum Genet 2007; 15: 831–836.
Choi YH, Cotterchio M, McKeown-Eyssen G et al: Penetrance of colorectal cancer among MLH1/MSH2 carriers participating in the colorectal cancer familial registry in Ontario. Hered Cancer Clin Pract 2009; 7: 14.
Bonadona V, Bonaiti B, Olschwang S et al: Cancer risks associated with germline mutations in MLH1, MSH2, and MSH6 genes in Lynch syndrome. JAMA 2011; 305: 2304–2310.
Antoniou AC, Chenevix-Trench G : Common genetic variants and cancer risk in Mendelian cancer syndromes. Curr Opin Genet Dev 2010; 20: 299–307.
Scott RJ, Lubinski J : Genetic epidemiology studies in hereditary non-polyposis colorectal cancer. Methods Mol Biol 2009; 472: 89–102.
Wijnen JT, Brohet RM, van Eijk R et al: Chromosome 8q23.3 and 11q23.1 variants modify colorectal cancer risk in Lynch syndrome. Gastroenterology 2009; 136: 131–137.
Talseth-Palmer BA, Brenne IS, Ashton KA et al: Colorectal cancer susceptibility loci on chromosome 8q23.3 and 11q23.1 as modifiers for disease expression in Lynch syndrome. J Med Genet 2010; 48: 279–284.
Houlle S, Charbonnier F, Houivet E et al: Evaluation of Lynch syndrome modifier genes in 748 MMR mutation carriers. Eur J Hum Genet 2011; 19: 887–892.
Londono-Vallejo JA : Telomere length heterogeneity and chromosome instability. Cancer Lett 2004; 212: 135–144.
Meeker AK, Hicks JL, Iacobuzio-Donahue CA et al: Telomere length abnormalities occur early in the initiation of epithelial carcinogenesis. Clin Cancer Res 2004; 10: 3317–3326.
Collins K, Mitchell JR : Telomerase in the human organism. Oncogene 2002; 21: 564–579.
Blasco MA, Lee HW, Hande MP et al: Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell 1997; 91: 25–34.
Calado RT, Chen J : Telomerase: not just for the elongation of telomeres. Bioessays 2006; 28: 109–112.
McKay JD, Hung RJ, Gaborieau V et al: Lung cancer susceptibility locus at 5p15.33. Nat Genet 2008; 40: 1404–1406.
Shete S, Hosking FJ, Robertson LB et al: Genome-wide association study identifies five susceptibility loci for glioma. Nat Genet 2009; 41: 899–904.
Wrensch M, Jenkins RB, Chang JS et al: Variants in the CDKN2B and RTEL1 regions are associated with high-grade glioma susceptibility. Nat Genet 2009; 41: 905–908.
Jin G, Xu L, Shu Y et al: Common genetic variants on 5p15.33 contribute to risk of lung adenocarcinoma in a Chinese population. Carcinogenesis 2009; 30: 987–990.
Landi MT, Chatterjee N, Yu K et al: A genome-wide association study of lung cancer identifies a region of chromosome 5p15 associated with risk for adenocarcinoma. Am J Hum Genet 2009; 85: 679–691.
Rafnar T, Sulem P, Stacey SN et al: Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat Genet 2009; 41: 221–227.
Petersen GM, Amundadottir L, Fuchs CS et al: A genome-wide association study identifies pancreatic cancer susceptibility loci on chromosomes 13q22.1, 1q32.1 and 5p15.33. Nat Genet 2010; 42: 224–228.
Hofer P, Baierl A, Feik E et al: MNS16A tandem repeats minisatellite of human telomerase gene: a risk factor for colorectal cancer. Carcinogenesis 2011; 32: 866–871.
Hosgood HD, Cawthon R, He X, Chanock S, Lan Q : Genetic variation in telomere maintenance genes, telomere length, and lung cancer susceptibility. Lung Cancer 2009; 66: 157–161.
Van Dyke AL, Cote ML, Wenzlaff AS et al: Chromosome 5p Region SNPs Are Associated with Risk of NSCLC among Women. J Cancer Epidemiol 2009; 2009: 242151.
Moreno V, Gemignani F, Landi S et al: Polymorphisms in genes of nucleotide and base excision repair: risk and prognosis of colorectal cancer. Clin Cancer Res 2006; 12: 2101–2108.
Pros E, Gomez C, Martin T, Fabregas P, Serra E, Lazaro C : Nature and mRNA effect of 282 different NF1 point mutations: focus on splicing alterations. Hum Mutat 2008; 29: E173–E193.
Messeguer X, Escudero R, Farre D, Nunez O, Martinez J, Alba MM : PROMO: detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002; 18: 333–334.
Farre D, Roset R, Huerta M et al: Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res 2003; 31: 3651–3653.
Cawthon RM : Telomere length measurement by a novel monochrome multiplex quantitative PCR method. Nucleic Acids Res 2009; 37: e21.
Mocellin S, Verdi D, Pooley KA et al: Telomerase reverse transcriptase locus polymorphisms and cancer risk: a field synopsis and meta-analysis. J Natl Cancer Inst 2012; 104: 840–854.
Ding Z, Green AG, Yang X, Chernenko G, Tang SC, Pater A : Retinoic acid inhibits telomerase activity and downregulates expression but does not affect splicing of hTERT: correlation with cell growth rate inhibition in an in vitro cervical carcinogenesis/multidrug-resistance model. Exp Cell Res 2002; 272: 185–191.
Pendino F, Dudognon C, Delhommeau F et al: Retinoic acid receptor alpha and retinoid-X receptor-specific agonists synergistically target telomerase expression and induce tumor cell death. Oncogene 2003; 22: 9142–9150.
Pickett HA, Baird DM, Hoff-Olsen P et al: Telomere instability detected in sporadic colon cancers, some showing mutations in a mismatch repair gene. Oncogene 2004; 23: 3434–3443.
Mendez-Bermudez A, Hills M, Pickett HA et al: Human telomeres that contain (CTAGGG)n repeats show replication dependent instability in somatic cells and the male germline. Nucleic Acids Res 2009; 37: 6225–6238.
Rampazzo E, Bertorelle R, Serra L et al: Relationship between telomere shortening, genetic instability, and site of tumour origin in colorectal cancers. Br J Cancer 2010; 102: 1300–1305.
Mendez-Bermudez A, Royle NJ : Deficiency in DNA mismatch repair increases the rate of telomere shortening in normal human cells. Hum Mutat 2011; 32: 939–946.
Djojosubroto MW, Choi YS, Lee HW, Rudolph KL : Telomeres and telomerase in aging, regeneration and cancer. Mol Cells 2003; 15: 164–175.
Burn J, Gerdes AM, Macrae F et al: Long-term effect of aspirin on cancer risk in carriers of hereditary colorectal cancer: an analysis from the CAPP2 randomised controlled trial. Lancet 2011; 378: 2081–2087.
Acknowledgements
We thank all the people responsible for genetic counseling and genetic testing in hereditary cancer at both Catalan Institute of Oncology and Leiden University Medical Center, and Gemma Aiza for technical support. This work was partly funded by the Spanish Ministry of Science and Innovation (grant BFU2009-10281 and Ramón y Cajal contract, both to LV; and fellowship to FB), the Scientific Foundation of Asociación Española Contra el Cáncer, the Carlos III Health Institute (ISCIIIRETIC: RD06/0020/1051 and RD06/0020/1050; FIS PI08/1635, FIS PI08/1359 and FIS PS09-01037; and fellowship to NS), the Catalan Health Institute and the Autonomous Government of Catalonia (2009SGR290, 2009SGR1489), and CIBERESP (CB07/02/2005).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on European Journal of Human Genetics website
Supplementary information
Rights and permissions
About this article
Cite this article
Bellido, F., Guinó, E., Jagmohan-Changur, S. et al. Genetic variant in the telomerase gene modifies cancer risk in Lynch syndrome. Eur J Hum Genet 21, 511–516 (2013). https://doi.org/10.1038/ejhg.2012.204
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejhg.2012.204
Keywords
This article is cited by
-
A genetic variant in telomerase reverse transcriptase (TERT) modifies cancer risk in Lynch syndrome patients harbouring pathogenic MSH2 variants
Scientific Reports (2021)
-
Correlation of human telomerase reverse transcriptase single nucleotide polymorphisms with in vitro fertilisation outcomes
Journal of Assisted Reproduction and Genetics (2019)
