- NEWS AND VIEWS
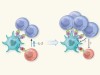
Peptide secretion triggers diabetes
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 560, 33-34 (2018)
doi: https://doi.org/10.1038/d41586-018-05710-z
References
Wan, X. et al. Nature 560, 107–111 (2018).
Banting, F. G. et al. Can. Med. Assoc. J. 12, 141–146 (1922).
Wan, X. & Unanue, E. R. Curr. Opin. Immunol. 46, 30–37 (2017).
Wallberg, M. & Cooke, A. Trends Immunol. 34, 583–591 (2013).
DiMeglio, L. A., Evans-Molina, C. & Oram, R. A. Lancet 391, 2449–2462 (2018).
Suri, A., Walters, J. J., Gross, M. L. & Unanue, E. R. J Clin. Invest. 115, 2268–2276 (2005).
Wan, X., Thomas, J. W. & Unanue, E. R. J. Exp. Med. 213, 967–978 (2016).
Middleton, D., Menchaca, L., Rood, H. & Komerofsky, R. Tissue Antigens 61, 403–407 (2003).
Pearson, J. A., Wong, F. S. & Wen, L. J. Autoimmun. 66, 76–88 (2016).
Livingstone, S. J. et al. J. Am. Med. Assoc. 313, 37–44 (2015).
 Read the paper: Pancreatic islets communicate with lymphoid tissues via exocytosis of insulin peptides
Read the paper: Pancreatic islets communicate with lymphoid tissues via exocytosis of insulin peptides
 The healthy diabetic cavefish conundrum
The healthy diabetic cavefish conundrum
 In the right place at the right time
In the right place at the right time