- NEWS AND VIEWS
Motion processing picks up speed in the brain
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 558, 38-39 (2018)
doi: https://doi.org/10.1038/d41586-018-04289-9
References
Hubel, D. H. & Wiesel, T. N. J. Physiol. 160, 106–154 (1962).
Lien, A. D. & Scanziani, M. Nature 558, 80–86 (2018).
Alonso, J.-M., Usrey, W. M. & Reid, R. C. J. Neurosci. 21, 4002–4015 (2001).
Ferster, D., Chung, S. & Wheat, H. Nature 380, 249–252 (1996).
Saul, A. B. & Humphrey, A. L. J. Neurophysiol. 64, 206–224 (1990).
Saul, A. B. & Humphrey, A. L. J. Neurophysiol. 68, 1190–1208 (1992).
Stanley, G. B. et al. J. Neurosci. 32, 9073–9088 (2012).
Reid, R. C., Soodak, R. E. & Shapley, R. M. J. Neurophysiol. 66, 505–529 (1991).
Livingstone, M. S. Neuron 20, 509–526 (1998).
McLean, J. & Palmer, L. A. Vision Res. 29, 675–679 (1989).
Lorente de No, R. in Physiology of the Nervous System (ed. Fulton, J.) 291–340 (Oxford Univ. Press, 1938).
Cruz-Martín, A. et al. Nature 507, 358–361 (2014).
Kremkow, J., Jin, J., Wang, Y. & Alonso, J. M. Nature 533, 52–57 (2016).
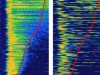 Read the paper: Cortical direction selectivity emerges at convergence of thalamic synapses
Read the paper: Cortical direction selectivity emerges at convergence of thalamic synapses
 These retinas are made for walkin’
These retinas are made for walkin’
 Species specific motion detectors
Species specific motion detectors