- NEWS AND VIEWS
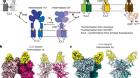
Force-activated ion channels in close-up
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 554, 469-470 (2018)
doi: https://doi.org/10.1038/d41586-018-01631-z
References
Murthy, S. E., Dubin, A. E. & Patapoutian, A. Nature Rev. Mol. Cell Biol. 18, 771–783 (2017).
Saotome, K. et al. Nature 554, 481–486 (2018).
Zhao, Q. et al. Nature 554, 487–492 (2018).
Guo, Y. R. & MacKinnon, R. eLife 6, e33660 (2017).
Coste, B. et al. Science 330, 55–60 (2010).
Ge, J. et al. Nature 527, 64–69 (2015).
Kellenberger, S. & Grutter, T. Mol. Biol. 427, 54–66 (2015).
Moroni, M., Servin-Vences, M. R., Fleischer, R. & Lewin, G. R. Preprint at bioRxiv http://dx.doi.org/10.1101/156489 (2017).
Wu, J., Goyal, R. & Grandl, J. Nature Commun. 7, 12939 (2016).
Kung, C. Nature 436, 647–654 (2005).
Brohawn, S. G. Ann. N. Y. Acad. Sci. 1352, 20–32 (2015).
Jin, P. et al. Nature 547, 118–122 (2017).
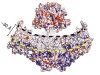 Read the paper: Structure of the mechanically activated ion channel Piezo1
Read the paper: Structure of the mechanically activated ion channel Piezo1
 Read the paper: Structure and mechanogating mechanism of the Piezo1 channel
Read the paper: Structure and mechanogating mechanism of the Piezo1 channel
 Stretched divisions
Stretched divisions