
Abstract
Within a multicellular tissue cells may coordinately form a singular or multiple polar axes, but it is unclear whether a common mechanism governs different types of polar axis formation. The phosphorylation status of PIN proteins, which is directly affected by the PINOID (PID) protein kinase and the PP2A protein phosphatase, is known to regulate the apical-basal polarity of PIN localization in bipolar cells of roots and shoot apices. Here, we provide evidence that the phosphorylation status-mediated PIN polarity switch is widely used to modulate cellular processes in Arabidopsis including multipolar pavement cells (PC) with interdigitated lobes and indentations. The degree of PC interdigitation was greatly reduced either when the FYPP1 gene, which encodes a PP2A called phytochrome-associated serine/threonine protein phosphatase, was knocked out or when the PID gene was overexpressed (35S::PID). These genetic modifications caused PIN1 localization to switch from lobe to indentation regions. The PP2A and PID mediated switching of PIN1 localization is strikingly similar to their regulation of the apical-basal polarity switch of PIN proteins in other cells. Our findings suggest a common mechanism for the regulation of PIN1 polarity formation, a fundamental cellular process that is crucial for pattern formation both at the tissue/organ and cellular levels.
Similar content being viewed by others

Introduction
In multicellular tissues, polarization or shape formation is coordinated among all cells within the population of a particular cell type so that they take the same polar or shape feature. For instance, in higher plants PIN proteins are polarized to the bottom end of cells in the root stele to direct the top-to-bottom auxin flow. In the leaf epidermis, all pavement cells (PCs) take the shape of interdigitated jigsaw puzzle pieces. However, it is unknown whether there is a unifying mechanism underlying the coordinated cell polarization or shape change during the development of multicellular tissues.
Cell intercalation is an important cellular mechanism for developmental patterning in multicellular organisms 1, 2, 3, 4, 5. Arabidopsis leaf PCs forming interdigitated lobes and indentations serve as a model system for the study of the mechanism underpinning cellular intercalation 6, 7, 8, 9, 10, 11. Our previous studies show that the interdigitated pattern of cell expansion in PCs involves two Rho GTPase-dependent pathways. The ROP2 GTPase is activated in the lobing region to promote lobe formation 6, 9. The ROP6 GTPase is localized in the indenting region to restrict outgrowth and promote indentation formation 9, 11. Mutual inhibition between ROP2 and ROP6 helps to establish alternating lobing and indenting regions along the plasma membrane (PM), generating a wave-like cell outline in PCs 9, 11. Both ROP2 and ROP6 pathways are activated by auxin through the cell surface auxin binding protein 1 (ABP1) 12. In the lobe region, auxin is transported to the cell surface by the PIN1 auxin exporter, which is localized to the lobe PM 12. By activating both ROP2 and ROP6, PIN1-exported auxin coordinates lobe formation with indentation promotion. Consequently PIN1 is polarly localized to the lobe PM in an interdigitated manner. However, the mechanism underlying the interdigitated PIN1 localization pattern is not clear.
PIN proteins localize to the same ends of bipolar cells to control directional auxin flow and auxin-gradient formation 13. It is known that protein phosphatase 2A (PP2A) and Ser/Thr protein kinase PINOID (PID) directly modulate the phophorylation status of PM-localized PINs and antagonistically mediate apical-basal polar targeting of PINs in roots and shoot apex in Arabidopsis 14, 15, 16, 17. In wild-type roots, PIN1 preferentially localizes to the basal end of stele cells to transport auxin from top to bottom. PIN1 hyper-phosphorylation induced by PID overexpression or PP2A downregulation caused PIN1 to switch from the basal to the apical side 14, 15, 16, 17. In the shoot apex, hypo-phosphorylated PIN proteins are switched from the apical to the basal side 16.
In this work, we demonstrate that the interdigitated pattern of PIN1 localization modulates interdigitated cell-expansion patterning in Arabidopsis leaf epidermal PCs and that PID- and PP2A-dependent phosphorylation status regulates PIN1 polarity switch between lobes and indentations, as in the apical-basal polarity switch in bipolar cells. This finding reveals a unifying mechanism for PIN-protein polarization that is involved in cell-cell coordination of cell expansion and auxin flow and auxin-gradient formation essential for tissue and organ patterning in plants.
Results
Genetic screen for mutations causing defects in interdigitated pattern of cell expansion in Arabidopsis PCs
To investigate mechanisms underlying the interdigitated pattern of PC expansion, we screened for T-DNA insertion mutations causing defective interdigitation (DID) in cotyledon PCs and isolated did1-1, which exhibited greatly reduced lobing at the early stage of PC development. We quantitatively analyzed the did-1 phenotype by measuring mean lobe numbers/area (lobes/μm2) in PCs of cotyledons 3 days after germination (Figure 1A) and the third true leaves (Figure 2A). In day 3 cotyledons, the did1-1 mutant had almost half of the lobe numbers (3.0 × 10−3/μm2) compared with wild type (5.6 × 10−3/μm2) (Figure 1B). Similar defects were found in true leaves (Figure 2A). In the third true leaves, wild-type and did-1 cells had 2.9 × 10−3 lobes/μm2 and 1.6 × 10−3 lobes/μm2, respectively (Figure 2B).

PC shape phenotype in fypp1 mutant and 35S::PID lines. (A) Representative PC shapes in the cotyledons from Col 0, fypp1-1, 35S::FYPP1 fypp1-1 and 35S::PID seedlings grown on agar medium 3 days after germination. White dotted outlines indicate the boundary of a single cell. Bars = 50 μm. (B) Quantitative analysis of mean lobe numbers in PCs of the third day cotyledon of Col 0, fypp1-1, fypp1-2, 35S::FYPP1 fypp1-1 and 35S::PID. Mean lobe numbers per area were measured as previously described 12. Both fypp1 alleles and 35S::PID show significantly fewer lobe numbers compared to Col 0 control (indicated by triple stars, P < 0.001, t-test). Mean lobe numbers in 35S::FYPP1 fypp1-1 were not significantly different from those of Col 0 (P > 0.01). (C) RT-PCR analysis of FYPP1 transcript levels in fypp1-1, fypp1-2 and 35S::FYPP1 fypp1-1. (D) T-DNA insertion positions of fypp1-1 and fypp1-2. The location of T-DNA insertion (443 bp after ATG) in fypp1-1 was analyzed by Tail-PCR and sequencing. The T-DNA insertion site at 5′ UTR in fypp1-2 (obtained from ABRC) was confirmed by PCR genotyping.

PC lobe formation in fypp1 and 35S::PID true leaves is insensitive to auxin treatment. (A) Representative images of PCs from the third true leaves of Col 0, fypp1-1, fypp1-2 and 35S::PID plants 2 days after treatment with MS liquid (DMSO control) and MS liquid containing 20 nM NAA. The outlines indicate typical cell shapes. Bars = 50 μm. (B) Quantitative analysis of mean lobe numbers in PCs of the third true leaves of Col 0, fypp1-1, fypp1-2 and 35S::PID with or without NAA treatment. NAA treatment increased mean lobe numbers/area significantly in Col 0 (P < 0.001, t-test) but not in fypp1-1, fypp1-2 and 35S::PID.
DID1 encodes the FYPP1 PP2A protein phosphatase
Tail-PCR analysis revealed that did1-1 contained a T-DNA insertion into the first intron (443 bp behind the ATG start codon) of the FYPP1 gene, which encodes a phytochrome-associated serine/threonine protein phosphatase 18. Another fypp1 insertion line containing a T-DNA inserted in the 5′ UTR, 135 bp front of the ATG start codon (Figure 1D), did1-2/fypp1-2, showed a similar phenotype (Figure 1A and B). RT-PCR analysis indicated fypp1 lines showed dramatic decrease in FYPP1 transcripts. Furthermore, the PC phenotype in did1-1/fypp1-1 was fully rescued by the 35S::FYPP1 construct stably introduced into the mutant (Figure 1A, 1B and 1C). The did1/fypp1 mutants also showed clearly shorter lobes in day 3 cotyledons, and this short-lobe phenotype was also rescued by 35S::FYPP1 (Supplementary information, Figure S1).
PCs in loss-of-function fypp1 mutants are similar to those in gain of PID function mutants
FYPP1/DID1 belongs to the PP2Ac family, whose members are catalytic subunits of the PP2A protein phosphatase complex. Members of the PP2Aa family, including PP2AA1, PP2AA2 and PP2AA3, which are regulatory subunits of the PP2A complex, have been shown to control the status of phosphorylation of PIN proteins 14. We found that PP2AA also affects pavement cell morphogenesis. The pp2aa1 pp2aa3 double mutant with strong defects in root development also shows a great decrease in lobe formation in PCs of cotyledons (2.035 × 10−3 lobes /μm2) compared with wild type (4.994 × 10−3 lobes /μm2) (Supplementary information, Figure S2A and S2B). PP2AA acts antagonistically with PID 19, 20, 21, 22 to modulate the phosphorylation status of PIN proteins and the polarity of their localization in roots and shoot apices 14, 15, 16, 17. Indeed a gain of PID function line (35S::PID) showed a defect in PC interdigitation similar to that in the did1-1 mutant (Figure 1A and 1B). Because PID- and PP2A-mediated PIN phosphorylation status regulates PIN polarity, these results imply that auxin transport may modulate the formation of the PC interdigitation pattern.
Exogenously applied auxin does not rescue PC lobe formation in fypp1 and 35S::PID
We considered two possible mechanisms by which FYPP1-mediated auxin transport may regulate PC interdigitation. FYPP1-mediated auxin transport could be required for the overall accumulation of auxin in leaves/cotyledons, which could act as a global signal to coordinate cell shape changes in the leaf epidermis. In this case, auxin transport-dependent auxin accumulation in cotelydons/leaves would be expected to occur prior to the formation of the jigsaw-puzzle PC shape. Alternatively, FYPP1 could mediate PIN1 localization to the tip of lobe, which has been implicated in the regulation of PC interdigitation 12. We reason that if FYPP1/PID-mediated auixn transport only affects global auxin accumulation in cotyledon/leaf, the PC interdigitation defect in the fypp1/did1 mutants and the 35S::PID lines should be rescued by exogenously applied auxin, as was the yuc1 yuc2 yuc4 yuc6 quadruple mutant defective in auxin biosynthesis 12. In contrast, we would predict that FYPP1 and PID directly affect PIN1 localization to the lobe PM if these lines fail to respond to exogenous auxin, as seen in loss of PIN function mutants 12. We treated the 3-week seedlings with 20-nM naphthalene-1-acetic acid (NAA, a synthetic auxin) in liquid MS medium. After 2-3 days of incubation, the third true leaves were stained with 5 μg/ml FM4-64 or FM1-43 for 1 h for visualization of PC shapes. Small protrusions were notably increased in wild-type PCs by 2 days after NAA treatment. The mean lobe numbers increased from 2.863 × 10−3 lobes/μm2 to 4.439 × 10−3 lobes/ μm2. In contrast, fypp1-1, fypp1-2 and 35S::PID mutants did not respond to this treatment (Figure 2A and 2B). Similar results were obtained with 3-day-old cotyledons (data not shown). Based on these results, we conclude that the direct cause of the interdigitation defect in the fypp1 mutants and the 35S::PID lines was not related to overall auxin accumulation in leaves that might be PIN dependent.
PIN1 localization is altered in 35S::PID and fypp1-1 PCs
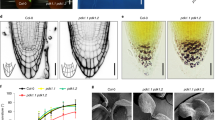
We next tested the hypothesis that FYPP1 regulates the polar subcellular localization of PIN1 to the lobe tip in PCs, which is critical for the interdigitated pattern of PC shapes 12. PIN1 is preferentially localized to the PM of lobe regions and thus displays an interdigitated pattern of localization along the PM of two neighboring cells bordered by the cell wall 12. To analyze the effect of PID/FYPP1-mediated phosphorylation 14, 16, 17 on PIN1 localization, we conducted PIN1 immunostaining in PCs of Col-0 wild-type control, 35S::PID and fypp1-1 leaves, and quantified regions of cells displaying PIN1 localization preferentially to lobes or indentations, or non-preferentially to both indentations and lobes (Figure 3A). The quantitative data showed that in wild-type PCs, 90% cell regions (73/81) had preferential PIN1 localization to lobes and only 10% cell regions displayed PIN1 localization to indentation regions (9%) or even PIN1 distribution to the PM (1%). In 35S::PID PCs, PIN1 localization was switched to the PM of indentation regions (Figure 3A). In 35S::PID leaves, 90% of cell regions displayed PIN1 localization to indentations, whereas 10% of cell regions showed PIN1 localization to lobes (4%) and even distribution to the PM (6%) (Figure 3B, 3C). In fypp1-1, PIN1 localization also lost the lobe preference and became preferentially localized to indentations as in 35S::PID cells, although a significant proportion of cells displayed even PIN1 distribution to the PM (Figure 3A). In this line, only 8% cell regions had PIN1 localized to lobes, while 92% of them showed PIN distribution evenly to the PM (41%) or preferentially to indentations (51%) (Figure 3B, 3C). These results suggest that PID kinase and PP2A phosphatase have opposite effects on PIN1 distribution between the lobe and indentation regions of PCs.

PIN1 localization is altered in 35S::PID and fypp1-1 PCs. (A) Subcellular localization of PIN1 was examined by using immunostaining with anti-PIN1 antibody as described in procedures. PIN1 was localized preferentially at the apical PM of lobes in Col 0 PCs, but in the PM of indentation regions in 35S::PID PCs. In fypp1-1 PCs, PIN1 was also shifted to the PM of indentation regions, but could sometimes be found in both lobe and indentation regions of the PM. Bars = 15 μm. (B) Schematic representation of PIN1 localization at the apical PM of lobes (1), indentation region of the PM (2) and both sides (3) of the cells. (C) Quantitative analysis of PIN1 localization patterns. The number indicates percentage of PM regions showing a particular localization pattern in PCs from three indicated lines: (1) PIN1 localization only to the lobe side, (2) PIN1 localization only to the indentation side and (3) PIN1 localization to both sides. The PIN1 localization pattern was quantified from more than 100 randomly chosen PCs.
PIN1 level affects PC shape
On the basis of the above observations, we hypothesize that PID and PP2A directly influence PIN1 localization patterns and subsequently the PC interdigitation pattern. This hypothesis is consistent with our previous observations that pin1 loss-of-function mutations induce defects in the PC interdigitation pattern 12 and that PC interdigitation patterning defect in pin1 and pp2a mutant could not be rescued by auxin (Figure 2; 12). To further test this hypothesis, we analyzed PC phenotype and PIN1 localization in PCs overexpressing PIN1. In 35S::PIN1 cotyledons, PCs displayed a dramatic increase in the number of lobes and a decrease in the size of lobes (Figure 4A, 4B). The number of lobes in 35S::PIN1 cotyledons (4.958 × 10−3 lobes/μm2) was nearly doubled compared to wild-type cotyledons (2.823 × 10−3 lobes/μm2) (Figure 4A and 4B). Furthermore, PIN1 localization in the 35S::PIN1 leaves was also altered. Compared to wild-type cells, the preference of PIN1 for the lobe region of the PM was much less pronounced (Figure 4C). In most cases, PIN1 tended to be more evenly distributed to the PM. The increased PM region containing PIN1 could explain the PC phenotype in 35S::PIN1 leaves (increased number of lobes with smaller lobe size).

PC interdigitation pattern and PIN1 localization are altered in 35S::PIN1 cotyledons. (A) Representative shapes of PCs from cotyledons of Col 0 and 35S::PIN1 seedlings 4 days after germination. Bars = 50 μm. (B) Quantitative analysis of lobe numbers in PCs of 4-day-old cotyledons in Col 0 and 35S::PIN1. 35S::PIN1 cotyledons shown in (A). 35S::PIN1 clearly increased PC lobe formation (P < 0.001, t-test). (C) PIN1 immunostaining in Col 0 and 35S::PIN1. In 35S::PIN1, PIN1 tended to accumulate to a high level in a large area of the PM within one PC. PIN1 was localized evenly to the PM at both opposing sides. White arrowheads show PIN1 localizations at indentation part, while white arrows indicate PIN1 localizations at lobing area. Bars = 15 μm.
Discussion
Our results indicate that a PID/PP2A-mediated signaling system is a universal mechanism for the regulation of PIN1 polarity in various cells regardless of the number of polarity axes in the cell or functional properties of the cell. Previous studies show that PID/PP2A modulates PIN1 polarity switch in bipolar cells involved in auxin flow and auxin-gradient formation in both root and shoot cells 14, 16. Similarly, in bipolar guard cells PID overexpression induced PIN1 localization to switch from the inner region of the PM to outer region (Supplementary information, Figure S3A and S3B, and Figure 5C). Interestingly, here we show that PID/PP2A regulates PIN1 polarity switch between regions of cells for outgrowth (lobe formation) and suppression of outgrowth (indent formation) in interdigitated PCs with multiple polar axes pointing at various directions.

Schematic view of the PID/PP2A-mediated PIN1 polarity switch in different cell types. (A) In PCs, PINOID kinase and FYPP1 phosphatase mediate reversal of the PIN1 polarity between lobes and indentations. We propose that hyper-phosphorylated and hypo-phosphorylated PIN1 is preferentially sorted for indentation and lobe tip targeting, respectively. Blue arrows represent PIN1 polarity switch. Red arrows show auxin transport direction. (B) In root cells, PINOID and PP2A mediate the apical-basal switch of PIN proteins. Hyper-phosphorylated PIN is preferentially targeted to the apical side, while hypo-phosphorylated PIN stays at the basal side. (C) PIN1 localization in guard cells. Hyper-phosphorylated PIN1 accumulates at the outer sides of the kidney-shaped guard cells, while hypo-phosphorylated PIN1 is targeted to the inner sides.
PID- and PP2A-mediated changes in PIN distribution in PCs are strikingly similar to PID- and PP2A-mediated PIN polarity switch between the basal and apical ends of root or shoot cells (Figure 5B). For example, PID overexpression or loss-of-function mutations in PP2AA (encoding a regulatory subunit of PP2A) caused PIN proteins to shift from the basal to the apical end of root cells 14, 16, 17, resembling the PIN1 polarity shift from lobes to indentations induced by 35S::PID or FYPP1 knockout. Recent reports show that PID and PP2A regulate the phosphorylation status of PIN proteins to control the polarity switch by affecting their directional endocytic recycling in these cells 14, 15, 16, 17, 23, 24. We propose that in PCs PID- and PP2A-modulated phosphorylation status similarly controls the PIN1 polarity switch between lobes and indentations (Figure 5A). The hypo-phosphorylated PIN1 (through the action of PP2A) is localized to the lobe region of the PM, while hyper-phosphorylated PIN1 (through the action of PID) is localized to the indented or non-lobing region of the PM. In wild-type cells, PID is likely to be rate-limiting, controlling a balanced level of PIN1 de-phosphorylation mediated by PP2A. In loss-of-PP2A or gain-of-PID cells, PIN1 phosphorylation level is tipped toward hyper-phosphorylation, causing PIN1 to localize to indentation or non-lobing regions. Our findings not only reveal a new mechanism for the control of the interdigitation patterning in PCs, i.e., PIN1 phosphorylation status modulates PIN1-dependent PC interdigitation patterns, but also demonstrate the conservation of this signaling mechanism in the regulation of different forms of cell polarity.
Given the significance of PIN polarity in plant pattern formation, the molecular mechanisms for PIN polarity control are important topics in cell and developmental biology, but are still poorly understood. The conservation of a PIN polarity switch mechanism in PCs opens up the possibility to use PCs for the study of this mechanism. This has clear advantages, as it has recently been shown that auxin promotes PIN1 localization to the lobe region of the PM through the auxin activation of the ABP1-ROP2 signaling pathway 12. This pathway promotes the accumulation of fine cortical actin microfilaments to the tip of lobes and inhibits the internalization of PIN1 9, 12. Downregulation of this pathway induced PIN1 internalization in PCs 12. Recent reports suggest that PID- and PP2A-mediated phosphorylation status of PIN proteins also regulates their endosomal trafficking to affect polarity switch in bipolar cells in root and shoot apices 14, 15, 16, 17, 23, 24. Furthermore, there are precedents suggesting that ROP signaling also regulates PIN localization in these cell types 25, 26, 27. Finally, ABP1 has been shown to affect PIN internalization in multipolar PCs as well as bipolar root cells 12, 28. These findings underscore the conservation of the mechanisms underlying PIN polarization between bipolar cells and interdigitated PCs. Evidence suggests that ROP signaling is regulated by the ABP1 auxin receptor, which appears to act at the cell surface 12, 28. PID-mediated PIN1 phosphorylation is also suggested to occur at the PM 23, 24. These observations raise an interesting possibility that ROP signaling and PID/PP2A-modulated PIN phosphorylation likely act coordinately or in the same signaling network to regulate PIN polarization. Therefore, our studies open up a new research direction in understanding how ROP signaling interfaces with the PID/PP2A-dependent phosphorylation switch in the control of PIN polarization, polar cell growth, auxin flow and auxin-gradient formation.
Materials and methods
Plant materials and growth conditions
Arabidopsis seeds were surface sterilized with 75% ethanol + 0.05% TritonX-100 for 10 min, sedimented and resuspended in 95% ethanol for 1-2 min. After ethanol was removed, seeds were allowed to dry in air briefly before being plated on MS (Murashige and Skoog) agar or sowed in soil. Following stratification at 4 °C for 3-4 days, they were incubated at 22 °C in a growth room under 16-h light/8-h dark cycles.
T-DNA insertion lines fypp1-2 (CS830707) was obtained from the Ohio State University Arabidopsis Biological Resource Center (ABRC). The 35S::PID line was previously described 16, 19. The pp2aa1 pp2aa3 double mutant was created by crossing pp2aa1 (rcn1 (root curling on NPA1)) to pp2aa3 (rcnL2) 29, 30.
Complementation of fypp1-1 by 35S::FYPP1
The coding sequence of FYPP1 gene (At1g50370) was amplified by RT-PCR using rTaq polymerase (TaKaRa) and FYPP1-specific primers, 5′-GGGGACAAGTTTGTACAAAAAAGCAGGCTTCatggatttagatcaatgga-3′ and 5′-GGGGACCACTTTGTACAAGAAAGCTGGGTTtcacaggaaataaggaac-3′, which contain attB recombination sites (Capital letters). The PCR product was cloned into pDONR (Invitrogen) by BP reaction and subsequently into the Gateway destination vector pH35GS (Riken Plant Science Center) using LR reactions. The FYPP1 sequence in the pDONR vector was confirmed by sequencing prior to subcloning into the destination vector. The final construct was introduced into Agrobacterium tumefaciens and transformed into the fypp1-1 mutant.
RT-PCR of analysis of FYPP1 expression
RNA was extracted using the RNeasy Plant Mini Kit (Qiagen), and was reverse-transcribed using SuperScript II First-Strand Synthesis System (Invitrogen). PCR amplification of the cDNA template was carried out by using FYPP1-specific primers: FYPP1-N-F 5′-GGTCATGTTCCCGAGACCAATTAC-3′ and FYPP1-N-R 5′-CACATTCCCACAACGGTAGCA-3′ (“N” means FYPP1 nonconserved domain). As a loading control, the same template cDNA was used for PCR amplification of the constitutively expressed ubiquitin gene involving two primers:
N-UBQ 5′-GGTGCTAAGAAGAGGAAGAAT-3′ and C-UBQ 5′-CTCCTTCTTTCTGGTAAACGT-3′ (“N” and “C” means N-terminal and C-terminal, respectively)
Microscopic analysis of PC shape
PC shapes of Arabidopsis cotyledons or true leaves were analyzed using confocal microscopy (Leica SP2 confocal microscope). The cell outlines were visualized using UV laser excitation (excitation: 351 nm or 364 nm, 50% laser power and emission: 400-600 nm) 12, because cell walls of these cells emit auto-fluorescence excitable by this wavelength. Alternatively, cell outlines were stained with 5 μg/ml FM4-64 (N-(3-triethylammoniumpropyl)-4-(6-(4-(diethylamino) phenyl)-hexatrienyl) pyridinium dibromide) dye or 5 μg/ml FM1-43 (N-(3-triethylammonium)propyl)-4-(4-(dibutylamino)styryl) pyridinium dibromide) and imaged using confocal microscopy with TRITC and FITC filters. Stack images (1.0- to 2.0-μm increments) were collected from more than 200 cells from five individual plants. Additional image quantification was conducted using MetaMorph software. Adobe Photoshop CS was used for text editing of all images.
Auxin treatment
NAA (Sigma) was dissolved in DMSO and prepared as a stock solution of 20 μM, which was added into liquid MS to obtain a final concentration of 20 nM for seedling treatments. Each treatment was repeated at least three times with three independent seedlings/line.
Immunofluorescence microscopy
PIN1 localization was examined by immunostaining following a previously described procedure 31. True leaves from 2-week-old seedlings were cut into small pieces about 2.5 mm2 and immediately submerged into a fixation buffer (PEMT (50mM PIPES PH = 6.8, 5mM EGTA, 1mM Mg2+, 0.05% TX-100) plus 1.5% formaldehyde) for 1 h. Fixed leaf tissues were washed in PEMT two to three times for 10 min each, blotted on slides, overlaid with cover glass, and plunged into liquid N2 until well frozen. Frozen tissues were shattered by applying pressure on a chilled smooth surface and washed into petri dish with cold permeabilization buffer (1× PBS, 1% TX-100) while tissue was still frozen. Shattered tissue pieces were incubated in room temperature for 2 h, transferred to wash buffer (1× PBS, 50mM glycine) in eppendorf tubes, and incubated for 30 min. The fixed tissues were incubated with the primary antibody (anti-PIN1, 1:1 000) 37 °C for 3 h and washed with the wash buffer three times for 10 min each. The FITC-conjugated anti-rabbit IgG (1:200, Sigma) secondary antibody was then added and incubated at 37 °C for 1-3 h. Stained tissues were washed with PBS three times for 10 min each and transferred into 0.1% (w/V) para-phenylene diamine in 1:1 PBS-glycerol. Following overnight incubation at 4 °C, stained tissues were observed under a Leica SP2 confocal microscope system using a 63× water lens. Stack images were taken by scanning at 0.5- to 1.0-μm increments. Lobe and indentation regions containing PIN1 were quantified from over 100 cells from three independent experiments. Preferential localization of PIN1 to lobes or indentation or equal localization to both regions was determined by eyeballing of the confocal stacked images.
References
McEwen DG, Peifer M . Wnt signaling: the naked truth? Curr Biol 2001; 11:R524–R526.
Goto T, Davidson L, Asashima M, Keller R . Planar cell polarity genes regulate polarized extracellular matrix deposition during frog gastrulation. Curr Biol 2005; 15:787–793.
Klein TJ, Mlodzik M . Planar cell polarization: an emerging model points in the right direction. Annu Rev Cell Dev Biol 2005; 21:155–176.
Settleman J . Intercalating Arabidopsis leaf cells: a jigsaw puzzle of lobes, necks, ROPs, and RICs. Cell 2005; 120:570–572.
Price MH, Roberts DM, McCartney BM, Jezuit E, Peifer M . Cytoskeletal dynamics and cell signaling during planar polarity establishment in the Drosophila embryonic denticle. J Cell Sci 2006; 119(Pt 3):403–415.
Fu Y, Li H, Yang Z . The ROP2 GTPase controls the formation of cortical fine F-actin and the early phase of directional cell expansion during Arabidopsis organogenesis. Plant Cell 2002; 14:777–794.
Smith LG . Cytoskeletal control of plant cell shape: getting the fine points. Curr Opin Plant Biol 2003; 6:63–73.
Li S, Blanchoin L, Yang Z, Lord EM . The putative Arabidopsis arp2/3 complex controls leaf cell morphogenesis. Plant Physiol 2003; 132:2034–2044.
Fu Y, Gu Y, Zheng Z, Wasteneys G, Yang Z . Arabidopsis interdigitating cell growth requires two antagonistic pathways with opposing action on cell morphogenesis. Cell 2005; 120:687–700.
Yang Z . Cell polarity signaling in Arabidopsis. Annu Rev Cell Dev Biol 2008; 24:551–575.
Fu Y, Xu T, Zhu L, Wen M, Yang Z . A ROP GTPase signaling pathway controls cortical microtubule ordering and cell expansion in Arabidopsis. Curr Biol 2009; 19:1827–1832.
Xu T, Wen M, Nagawa S, et al. Cell surface- and rho GTPase-based auxin signaling controls cellular interdigitation in Arabidopsis. Cell 2010; 143:99–110.
Wisniewska J, Xu J, Seifertova D, et al. Polar PIN localization directs auxin flow in plants. Science 2006; 312:883.
Michniewicz M, Zago MK, Abas L, et al. Antagonistic regulation of PIN phosphorylation by PP2A and PINOID directs auxin flux. Cell 2007; 130:1044–1056.
Huang F, Zago MK, Abas L, et al. Phosphorylation of conserved PIN motifs directs Arabidopsis PIN1 polarity and auxin transport. Plant Cell 2010; 22:1129–1142.
Friml J, Yang X, Michniewicz M, et al. A PINOID-dependent binary switch in apical-basal PIN polar targeting directs auxin efflux. Science 2004; 306:862–865.
Kleine-Vehn J, Huang F, Naramoto S, et al. PIN auxin efflux carrier polarity is regulated by PINOID kinase-mediated recruitment into GNOM-independent trafficking in Arabidopsis. Plant Cell 2009; 21:3839–3849.
Kim DH, Kang JG, Yang SS, et al. A phytochrome-associated protein phosphatase 2A modulates light signals in flowering time control in Arabidopsis. Plant Cell 2002; 14:3043–3056.
Benjamins R, Quint A, Weijers D, Hooykaas P, Offringa R . The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 2001; 128:4057–4067.
Christensen SK, Dagenais N, Chory J, Weigel D . Regulation of auxin response by the protein kinase PINOID. Cell 2000; 100:469–478.
Sukumar P, Edwards KS, Rahman A, Delong A, Muday GK . PINOID kinase regulates root gravitropism through modulation of PIN2-dependent basipetal auxin transport in Arabidopsis. Plant Physiol 2009; 150:722–735.
Zourelidou M, Muller I, Willige BC, et al. The polarly localized D6 PROTEIN KINASE is required for efficient auxin transport in Arabidopsis thaliana. Development 2009; 136:627–636.
Zhang J, Nodzynski T, Pencik A, Rolcik J, Friml J . PIN phosphorylation is sufficient to mediate PIN polarity and direct auxin transport. Proc Natl Acad Sci USA 2010; 107:918–922.
Dhonukshe P, Huang F, Galvan-Ampudia CS, et al. Plasma membrane-bound AGC3 kinases phosphorylate PIN auxin carriers at TPRXS(N/S) motifs to direct apical PIN recycling. Development 2010; 137:3245–3255.
Jaillais Y, Santambrogio M, Rozier F, et al. The retromer protein VPS29 links cell polarity and organ initiation in plants. Cell 2007; 130:1057–1070.
Hazak O, Bloch D, Poraty L, et al. A rho scaffold integrates the secretory system with feedback mechanisms in regulation of auxin distribution. PLoS Biol 2010; 8:e1000282.
Li L, Xu J, Xu ZH, Xue HW . Brassinosteroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis. Plant Cell 2005; 17:2738–2753.
Robert S, Kleine-Vehn J, Barbez E, et al. ABP1 mediates auxin inhibition of clathrin-dependent endocytosis in Arabidopsis. Cell 2010; 143:111–121.
Garbers C, DeLong A, Deruere J, Bernasconi P, Soll D . A mutation in protein phosphatase 2A regulatory subunit A affects auxin transport in Arabidopsis. EMBO J 1996; 15:2115–2124.
Rashotte AM, DeLong A, Muday GK . Genetic and chemical reductions in protein phosphatase activity alter auxin transport, gravity response, and lateral root growth. Plant Cell 2001; 13:1683–1697.
Wasteneys GO, Willingale-Theune J, Menzel D . Freeze shattering: a simple and effective method for permeabilizing higher plant cell walls. J Microsc 1997; 188 (Part 1):51–61.
Acknowledgements
We are grateful to members of the Yang group for stimulating discussions during the course of this work. This work is supported by grants from the US National Institute of General Medical Sciences (GM081451 and GM081451-03S2). HL was partly supported by Chinese Scholarship Council. PD was supported by Netherlands Scientific Organization NWO-VENI grant and BS by NWO Spinoza prize.
Author information
Authors and Affiliations
Corresponding author
Additional information
( Supplementary information is linked to the online version of the paper on the Cell Research website.)
Supplementary information
Supplementary information, Figure S1
Short lobe phenotype of fypp1 can be rescued by 35S::FYPP1. (PDF 625 kb)
Supplementary information, Figure S2
Pavement cell shapes of cotyledons from WT and pp2aa1 pp2aa3 double mutant. (PDF 433 kb)
Supplementary information, Figure S3
PIN1 localization in guard cells was also switched in 35S::PID plants. (PDF 151 kb)
Rights and permissions
About this article
Cite this article
Li, H., Lin, D., Dhonukshe, P. et al. Phosphorylation switch modulates the interdigitated pattern of PIN1 localization and cell expansion in Arabidopsis leaf epidermis. Cell Res 21, 970–978 (2011). https://doi.org/10.1038/cr.2011.49
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cr.2011.49
Keywords
This article is cited by
-
Involvement of differentially accumulated proteins and endogenous auxin in adventitious root formation in micropropagated shoot cuttings of Cedrela fissilis Vellozo (Meliaceae)
Plant Cell, Tissue and Organ Culture (PCTOC) (2022)
-
Auxin-induced signaling protein nanoclustering contributes to cell polarity formation
Nature Communications (2020)
-
Mining and expression analysis of candidate genes involved in regulating the chilling requirement fulfillment of Paeonia lactiflora ‘Hang Baishao’
BMC Plant Biology (2017)
-
Cytokinin signaling regulates pavement cell morphogenesis in Arabidopsis
Cell Research (2013)
