
Abstract
Sericulture has been greatly advanced by applying hybrid breeding techniques to the domesticated silkworm, Bombyx mori, but has reached a plateau during the last decades. For the first time, we report improved silk yield in a GAL4/UAS transgenic silkworm. Overexpression of the Ras1CA oncogene specifically in the posterior silk gland improved fibroin production and silk yield by 60%, while increasing food consumption by only 20%. Ras activation by Ras1CA overexpression in the posterior silk gland enhanced phosphorylation levels of Ras downstream effector proteins, up-regulated fibroin mRNA levels, increased total DNA content, and stimulated endoreplication. Moreover, Ras1 activation increased cell and nuclei sizes, enriched subcellular organelles related to protein synthesis, and stimulated ribosome biogenesis for mRNA translation. We conclude that Ras1 activation increases cell size and protein synthesis in the posterior silk gland, leading to silk yield improvement.
Similar content being viewed by others

Introduction
Hybrid breeding deliberately crosses closely or distantly related species to produce new lines with desirable properties. This conventional technique has been widely utilized to improve the yield of a variety of cultured plants and animals. Although hybrid silkworm breeding has greatly benefited sericulture in the past, during the last four decades, sericulture has reached a plateau, mostly due to the inherent threshold of this technique. To break through the bottleneck of silk production, new techniques, such as molecular breeding, must be developed for the domesticated silkworm, Bombyx mori. Molecular breeding consists of two modern breeding strategies, namely marker-assisted selection (MAS) and marker-assisted backcrossing (MABC). Transgenic breeding is the most important MABC technique 1 and is thought to possess great potential for improving silk production.
The Bombyx silk gland is composed of three regions: the anterior, middle and posterior silk glands. The anterior silk gland is responsible for silk spinning, the middle silk gland secretes sericin, and the posterior silk gland produces fibroin. Silk fiber is made of fibroin, and fibroin production by the posterior silk gland is directly proportional to silk yield. Fibroin production is determined by two factors: gland size and fibroin protein synthesis in the posterior silk gland 2, 3, 4, 5, 6. The idea that increasing posterior silk gland size and/or stimulating fibroin protein synthesis by transgenic breeding could improve fibroin production and thus improve silk yield is very attractive, although to date no such attempt has been reported.
The Ras oncogene encodes a small GTPase involved in both normal development and aberrant biological processes, such as tumorigenesis and developmental disorders. Ras activation enables high affinity interactions with its downstream effector proteins, including Raf and PI3K110. The Raf-MAPK and PI3K-Akt-TORC1-S6K/4EBP pathways participate in a variety of cellular and molecular events, particularly those related to cell growth and protein synthesis 7. For example, RasCA overexpression in the prothoracic gland in the fruitfly, Drosophila melanogaster, significantly reduces body size by increasing cell size in the gland and promoting ecdysone production 8.
There are three Ras genes in the Bombyx genome 9, 10, but their biological significance is largely unknown. Because the posterior silk gland grows dramatically during larval life and its protein synthesis capacity is extremely high at the last instar stage 2, 3, we hypothesized that Ras activity might be involved in regulating fibroin production and silk yield. Here, we report that, using the binary GAL4/UAS transgenic system 11, 12, overexpression of the Ras1CA oncogene specifically in the Bombyx posterior silk gland increases cell size and protein synthesis in the posterior silk gland and leads to silk yield improvement. Combining properties of both hybrid and molecular breeding techniques in Bombyx may produce a new breakthrough in sericulture.
Results and Discussion
Endogenous Ras activity correlates with posterior silk gland growth
We first investigated whether Ras activity is involved in fibroin production and silk yield. Phosphorylation levels of MAPK, a general readout for Ras activity 7, in the posterior silk gland were measured daily from day 2 of the 4th instar through the prepupal stage. MAPK phosphorylation was undetectable during the early 4th instar stage, gradually increased from the 4th larval molt to day 4 of the 5th instar, remained at high levels until the wandering stage, and rapidly decreased during pupation (Supplementary information, Figure S1A). The pattern of MAPK phosphorylation during development correlated with and slightly preceded the growth of the posterior silk gland (Supplementary information, Figure S1B). The developmental profile of Akt phosphorylation in the posterior silk gland was also measured. Unlike MAPK, Akt phosphorylation did not significantly correlate with the growth rate of the posterior silk gland (Supplementary information, Figure S2A). The reason could be that Akt phosphorylation results from multiple signal transduction pathways in addition to the Ras-PI3K110 pathway 7, 13. We then injected the PI3K110 inhibitor wortmannin into larvae on day 1 of the 5th instar. Wortmannin treatment reduced silk gland size, decreased whole body weight, and delayed larval development (Supplementary information, Figure S2B-S2E). From the above experimental data, we infer that Ras activity positively regulates fibroin production in the posterior silk gland.
Ras1CA overexpression improves silk yield
Approximately 10 years ago, a stable germ-line transformation method was established for the silkworm using the transposable element piggyBac as the transgenic vector 14. Based on this method, a binary GAL4/UAS system for targeted gene expression was developed 11 and was further applied to analyze gene function of juvenile hormone esterase 12. A major advantage of the GAL4/UAS system is its ability to target gene expression in a tissue- or stage-specific manner 15.
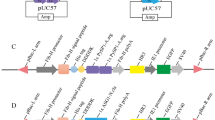
In Bombyx, the Ras1 mutant Ras1V12 (termed Ras1CA) is constitutively active 7, 9. To improve silk yield by transgenic breeding, the GAL4/UAS system was used to overexpress Ras1CA specifically in the posterior silk gland (Figure 1). The resulting transgenic silkworms, Fil-GAL4/UAS-Ras1CA [D(+)E(+)], showed an increase in posterior silk gland size of >60% compared to controls: wild type [D(−)E(−)], Fil-GAL4 [D(+)E(−)], and UAS-Ras1CA [D(−)E(+)], with ∼100% size increase in females (Figure 2A-A”) and ∼40% size increase in males (Supplementary information, Figure S3A). Conversely, tissue weights of the middle silk gland in the 4 groups were nearly identical. The cocoon weight in [D(+)E(+)] silkworms increased ∼40% compared to controls (Figure 2B and 2B'; Supplementary information, Figure S3B), which was a smaller proportional change than the posterior silk gland weight (60%). This is because the cocoon is composed of both sericin, produced by the middle silk gland, and fibroin, produced by the posterior silk gland. Interestingly, although the larval body weight of [D(+)E(+)] silkworms increased ∼20% (Figure 2C; Supplementary information, Figure S3C), the pupal body weight did not change (Figure 2D; Supplementary information, Figure S3D). We infer that the increased larval body weight was due to the increased posterior silk gland size.

The binary GAL4/UAS transgenic silkworm system and 4 different phenotypes. (A) The GAL4 (pBac{Fil-GAL4-3XP3-DsRed}) and UAS (pBac{UAS-Ras1v12-3XP3-EGFP}) plasmid constructs. (B) The UAS lines are crossed with the GAL4 lines to generate the Fil-GAL4/UAS-Ras1CA lines, in which Ras1CA was specifically overexpressed in the posterior silk gland. The progeny of this crossing shows four different phenotypes in terms of eye color (1) neither DsRed2-positive nor EGFP-positive, wild type, [D(−)E(−)]; (2) only DsRed2-positive, GAL4 lines, [D(+)E(−)]; (3) only EGFP-positive, UAS lines, [D(−)E(+)]; and (4) both DsRed2-positive and EGFP-positive, GAL4/UAS lines, [D(+)E(+)].

Increasing posterior silk gland size by Ras1CA overexpression greatly improves silk yield but has relatively little effects on food consumption. Three biological replicates are conducted and results from one replicate are presented. Mean ± SEM, n ≥ 25. The bars labeled with different lowercase letters are significantly different (P < 0.05, ANOVA). Data from females are shown. (A-A”) In comparison with the control silkworms: wild type [D(−)E(−)], Fil-GAL4 [D(+)E(−)], and UAS-Ras1CA [D(−)E(+)], posterior silk gland size in the transgenic silkworm Fil-GAL4/UAS-Ras1CA [D(+)E(+)] is significantly increased. (A) The anterior silk gland and the middle silk gland sizes from [D(+)E(+)] are not altered but its posterior silk gland size is increased. Blue arrow points to the area separating the anterior silk gland and the middle silk gland and green arrow points to the area separating the middle silk gland and the posterior silk gland. (A' and A”) A comparison of the posterior silk gland. (B and B') The cocoon weight of female [D(+)E(+)] is increased. (C) The larval body weight of female [D(+)E(+)] is increased. (D) The pupal body weight of female [D(+)E(+)] is not altered. It takes [D(+)E(+)] > 6 more hours to complete the feeding stage of the 5th instar. Food consumption by [D(+)E(+)] is increased.
[D(+)E(+)] silkworms required ∼6 more hours to complete the feeding stage of the 5th instar than the control silkworms (Figure 2E). Consequently, [D(+)E(+)] silkworms ate ∼20% more mulberry leaves during the 5th instar (Figure 2F). Considering a 60% increase in silk yield and only a 20% increase of food consumption, we estimate an approximate 30% improvement in the efficiency of converting food into silk in [D(+)E(+)] silkworms, indicating that the additional food was mostly converted into increased posterior silk gland size and thus silk yield. These findings represent a proof of principle that the transgenic silkworm Fil-GAL4/UAS-Ras1CA, which produces more silk but consumes only moderately more food, has potential in sericulture.
Ras activation stimulates Ras downstream effectors
We reasoned that increasing posterior silk gland size in [D(+)E(+)] silkworms was due to Ras1CA overexpression and thus Ras activation. Quantitative real-time PCR (qPCR) showed that Ras1 mRNA level in the posterior silk gland of [D(+)E(+)] silkworm was ∼10-fold higher than in controls (Figure 3A). In consequence, Ras activity was significantly increased (Figure 3B), as detected by a Ras Activation Assay Kit that consists of a GST fusion protein corresponding to the human Ras binding domain of Raf-1. Moreover, Ras1 activation enhanced the phosphorylation levels of MAPK, Akt, S6K, and 4EBP. Unlike the Ras downstream effectors, phosphorylation of InR 16, which is not a Ras downstream effector but acts upstream of the Ras-Raf-MAPK and the Ras-PI3K-Akt-TORC1-S6K/4EBP pathways 13, was not stimulated by Ras1 activation (Figure 3C). These results indicate that the Ras downstream effector proteins, Raf and PI3K110, were directly activated by Ras1CA overexpression in the posterior silk gland. In contrast to the posterior silk gland, in the middle silk gland, neither the Ras1 mRNA levels nor the phosphorylation levels of the Ras downstream effector proteins exhibited any significant differences between the four groups (data not shown), confirming that Fil-GAL4 specifically targets gene expression in the posterior silk gland 11. Taken together, we conclude that Ras1CA overexpression in the posterior silk gland increases Ras activity and stimulates Ras downstream effectors in the transgenic silkworm Fil-GAL4/UAS-Ras1CA.

Ras1CA overexpression in the posterior silk gland increases Ras1 activity and stimulates phosphorylation of Ras downstream effectors as well as RpS6 protein level. Three biological replicates are conducted and results from one replicate are presented. Mean ± SEM. Five males and five females are used in each replicate. (A) Ras1 mRNA level in the posterior silk gland from [D(+)E(+)] is ∼10-fold higher than those from the control silkworms [D(−)E(−)], [D(+)E(−)] and [D(−)E(+)]. The bars labeled with different lowercase letters are significantly different (P < 0.05, ANOVA). (B) Ras activity in the posterior silk gland of [D(+)E(+)] is increased. (C) Phosphorylation levels of MAPK, Akt, S6K and 4EBP as well as RpS6 protein level, but not InR phosphorylation level in the posterior silk gland from [D(+)E(+)] is increased. Tubulin is used as a loading control.
Ras activation increases fibroin mRNA levels and total DNA content
We then determined whether Ras activation in the posterior silk gland up-regulated fibroin mRNA levels in the transgenic silkworm Fil-GAL4/UAS-Ras1CA. As determined by qPCR, the mRNA levels of three major fibroin genes, fibroin heavy chain (Fih), Fil, and p25 in the posterior silk gland from [D(+)E(+)] silkworms were all up-regulated 2- to 3-fold compared to controls (Figure 4A-4C). The mRNA level of the transcription factor gene, bHLH, which is expressed exclusively in the silk gland and regulates the fibroin genes 17, 18, was ∼3-fold higher in [D(+)E(+)] silkworms than in controls (Figure 4D). The mechanism by which Ras activation up-regulates bHLH and the fibroin genes is unclear and merits further investigation.

Ras1 activation in the posterior silk gland up-regulates fibroin protein mRNA levels, increases total DNA content, and stimulates endoreplication. Three biological replicates are conducted and results from one replicate are presented. Mean ± SEM. Five males and five females are used in each replicate. The bars labeled with different lowercase letters are significantly different (P < 0.05, ANOVA). (A-D) The mRNA levels of three major fibroin proteins genes, fibroin heavy chain (Fih) (A), fibroin light chain (Fil) (B) and p25 (C) as well as a transcription factor gene bHLH (D) in the posterior silk gland from [D(+)E(+)] are 2-3-fold in comparison with those in the control silkworms [D(−)E(−)], [D(+)E(−)] and [D(−)E(+)]. (E and F) Total DNA content (E) and the mRNA levels of Cyclin E (CycE) and Cyclin D (CycD) (F) in the posterior silk gland from [D(+)E(+)] are 2-3-fold in comparison with those in the controls.
The silk gland is a typical endoreplicative tissue 19. Its endoreplicative cycles, which can be estimated by total DNA content, directly reflect silk yield 4. As compared to controls, total DNA content in the posterior silk gland of [D(+)E(+)] silkworms was nearly doubled (Figure 4E), implying that Ras activation promotes the endoreplicative cycles. The Raf-MAPK pathway plays a critical role in controlling cell proliferation. MAPK consists of 3 subfamilies, ERK, JNK and P38. Activated MAPK translocates to the nucleus and phosphorylates transcription factors, such as Myc, ELK, and Jun. These transcription factors up-regulate the Cyclin genes, which stimulate the G1 to S phase progression in cell cycle 7, 20. In parallel, Signaling through TORC1-S6K/4EBP also activates Myc to up-regulate the Cyclin genes 21. In Drosophila, Cyclin E and Cyclin D drive the endoreplication of larval tissues 22, 23. Consistent with the doubled DNA content, the mRNA levels of Cyclin E and Cyclin D in the [D(+)E(+)] posterior silk gland were ∼3-fold higher than in controls (Figure 4F). To confirm that the endoreplicative cycles of the posterior silk gland were stimulated in [D(+)E(+)] silkworms, we examined endoreplicative activity by BrdU labeling at the early wandering stage. BrdU labeling in the posterior silk gland of [D(+)E(+)] silkworms was much stronger than in controls (Supplementary information, Figure S4). As shown in Figure 2E, ∼6 more hours were required for [D(+)E(+)] silkworms to complete the feeding stage of the 5th instar compared to controls, suggesting that the additional 6 hours might provide enough time for posterior silk gland of the transgenic silkworms to undergo an additional endoreplicative cycle. These results demonstrate that Ras activation up-regulates Cyclin E and Cyclin D, promotes endoreplicative cycles, and increases total DNA content in the posterior silk gland of the transgenic silkworm Fil-GAL4/UAS-Ras1CA.
Ras activation increases cell size and protein synthesis
The final number of cells in the Bombyx silk gland is determined at an early embryonic stage. However, cell size increases during late embryonic development and continues increasing throughout all larval stages 3, 19, indicating that the silkworm posterior silk gland size is determined by cell size rather than cell number. Because Fil-GAL4 11 is specifically expressed in the posterior silk gland during late larval stages, the increased posterior silk gland size caused by Ras activation should be due to increased cell size. Although not yet well-characterized in Bombyx, cell size control is well understood in Drosophila. The evolutionarily conserved insulin/insulin-like growth factor signaling pathway (IIS) in Drosophila plays a crucial role in controlling nutrient-dependent cell growth 13, 24. In addition to the canonical IIS pathway, the TORC1 pathway promotes cell growth through its action on translational initiation, ribosome biogenesis, and nutrient storage 25, 26. Although little is known about the IIS-TORC1 pathways in Bombyx 16, 27, Ras activation significantly enhanced the phosphorylation levels of Akt, S6K and 4EBP (Figure 3C) and positively affected cell size in the posterior silk gland (Figure 2A; Supplementary information, Figure S3). Optical microscopy revealed that cell size in the posterior silk gland of [D(+)E(+)] silkworms was significantly increased with enlarged nuclei (Figure 5A and 5A'; Supplementary information, Figure S5). We then examined ultrastructural changes in the posterior silk gland cells of [D(+)E(+)] silkworms using transmission electron microscopy. Similar to the optical microscopy results, the nuclei were enlarged but maintained regular morphology (Figure 5B and 5B'). Moreover, subcellular organelles related to protein synthesis, including the rough endoplasmic reticula, mitochondria and secretory vesicles, were significantly enriched (Figure 5C-5D). We conclude that cell size and protein synthesis are enhanced in the posterior silk gland of the transgenic silkworm Fil-GAL4/UAS-Ras1CA.

Ras1 activation in the posterior silk gland increases cell and nucleolus size and enriches subcellular organelles related to protein synthesis. (A and A') Optical microscopy observations of the posterior silk gland. Cell size and nuclei size (pointed by arrows) in the posterior silk gland of [D(+)E(+)] (A') was increased in comparison with [D(−)E(−)] (A). Females are used in these experiments. (B-C') Transmission electron microscopy observations of the posterior silk gland cell. The nuclei size of posterior silk gland cell from [D(+)E(+)] (B') is enlarged with regular morphology compared to [D(−)E(−)] (B). Subcellular organelles related to protein synthesis, including layer rough endoplasmic reticula (RER, short arrows), mitochondria (middle arrows) and secretory vesicles (long arrows) are enriched in the posterior silk gland cell of [D(+)E(+)] (C') in comparison with [D(−)E(−)] (C). (D) A statistic comparison of RER, mitochondria and secretory vesicles in the posterior silk gland cell of [D(+)E(+)] and [D(−)E(−)]. Mean ± SEM. The bars labeled with different lowercase letters are significantly different (P < 0.05, ANOVA).
Ras activation increases ribosome biogenesis
The Ras-stimulated proteins TORC1 and Myc play a crucial role in the regulation of protein synthesis, partially by up-regulating ribosome biosynthesis genes and thus promoting ribosome biogenesis 13, 20, 24, 25, 26. The enrichment of rough endoplasmic reticula in [D(+)E(+)] silkworms (Figure 5C and 5C') indicates increased ribosome biogenesis. qPCR revealed that the mRNA levels of several ribosomal protein genes, including mRpL1, RpL12, RpS13, and RpS6, were all increased 2- to 3-fold in the posterior silk gland of [D(+)E(+)] silkworms at the early wandering stage (Figure 6). Moreover, western blot analysis showed that the RpS6 protein level was significantly increased (Figure 3C, RpS6). These data demonstrate that, consistent with the enrichment of subcellular organelles related to protein synthesis (Figure 5C-5D), Ras1 activation stimulates ribosome biogenesis for mRNA translation. These changes increase the protein synthesis ability in the posterior silk gland of the transgenic silkworm Fil-GAL4/UAS-Ras1CA.

Ras1 activation in the posterior silk gland up-regulates ribosomal protein genes. The mRNA levels of 4 ribosomal protein genes, including mRpl1 (A), Rpl12 (B), RpS13 (C) and RpS6 (D) in the posterior silk gland from [D(+)E(+)] are 2-3-fold in comparison with those in the control silkworms [D(−)E(−)], [D(+)E(−)] and [D(−)E(+)].Three biological replicates are conducted and results from one replicate are presented. Mean ± STEDV. Five males and five females are used in each replicate. The bars labeled with different lowercase letters are significantly different (P < 0.05, ANOVA).
Based on the experimental data, we propose a hypothetical mechanism by which silk yield is improved in the transgenic silkworm Fil-GAL4/UAS-Ras1CA(Supplementary information, Figure S6). First, Ras1CA overexpression increases Ras activity, resulting in phosphorylation of Ras downstream effector proteins, Raf and PI3K110, which in turn activate the Raf-MAPK and PI3K-Akt-TORC1-S6K/4EBP pathways (Figure 3). Second, the activated Raf-MAPK and PI3K-Akt-TORC1-S6K/4EBP pathways up-regulate Cyclin E and Cyclin D (Figure 4F), which promote endoreplicative cycles (Supplementary information, Figure S4) and increase total DNA content (Figure 4E). The activated PI3K-Akt-TORC1-S6K/4EBP pathway also increases cell size (Figure 5A and 5A'; Supplementary information, Figure S5) by stimulating translation initiation, ribosome biogenesis, and nutrient storage (Figures 5C-5D and 6). The enlarged cell size and increased endoreplicative cycles lead to enhanced posterior silk gland size. Third, the activated PI3K-Akt-TORC1-S6K/4EBP pathway stimulates ribosome biogenesis for mRNA translation, resulting in enhanced protein synthesis ability (Figures 5C-5D and 6). Via an unknown mechanism, Ras activation up-regulates bHLH and the fibroin protein genes (Figure 4A-4D). Stimulated protein synthesis capacity and up-regulated fibroin mRNA levels lead to increased fibroin protein synthesis. Last, the increases in the size of the posterior silk gland and fibroin protein synthesis in the posterior silk gland ultimately result in increased fibroin production and thus silk yield (Figure 2; Supplementary information, Figure S3).
To our knowledge, this is the first report of a beneficial utilization of aberrant tissue development caused by Ras overexpression. We conclude that the transgenic silkworm Fil-GAL4/UAS-Ras1CA has potential for use in sericulture. Because the laboratory strain Dazao was used for the transgenic silkworm studies, we were not able to investigate any changes of silk quality. For industrial silk production, transgenic breeding should be performed in hybrid silkworm strains, which are normally used in the sericultural industry. Combining properties of both hybrid and molecular breeding techniques in Bombyx may produce a new breakthrough in sericulture.
Materials and Methods
Insects
The Bombyx strain, Dazao, was used for germ-line transformation. After plasmid and helper DNA was injected into preblastoderm eggs, the embryos were incubated at 25 °C with 95%-100% humidity until hatching. The larvae were reared on fresh mulberry leaves in the laboratory at 25 °C under 14 h light/10 h dark cycles 16.
The binary GAL4/UAS transgenic silkworm system
The pBac{Fil-GAL4-3XP3-DsRed} (Fil-GAL4) and pBac{UAS-Ras1v12-3XP3-EGFP} (UAS-Ras1CA) plasmid constructs (Supplementary information, Figure 3A) were generated as described below with the transposable element piggyBac as the transgenic vector 14. The 740-bp promoter region of the fibroin light chain (Fil) gene 11, 28 was amplified using forward and reverse primers containing AscI and BamHI sites, respectively. The BamHI-SacII fragment, which contained the GAL4 gene and Dmhsp70 terminator, was amplified from phsGAL4 29. The above two PCR fragments were conjugated, digested with AscI and SacII, and inserted into pBac{3XP3-DsRed} 14 to generate Fil-GAL4. The cDNA of Bombyx Ras1 oncogene 9 was amplified using forward primer containing EcoRI and reverse primer containing NotI. Ras1CA was generated by substituting valine for glycine at the 12th amino acid residue of Ras1 (Ras1V12), which impairs the protein's ability to hydrolyze GTP 7. The HindIII-EcoRI fragment of the UAS sequence from pUAST 29 was inserted into pEGFP-N1 (Clontec), which contained the EGFP gene and SV40 terminator, to form an intermediate construct. The EGFP gene in the construct was replaced by the EcoRI-NotI Ras1CA fragment to generate pUAS-Ras1CA. The pUAS-Ras1CA fragment was then digested with NheI and AflII and inserted into pBac{3XP3-EGFP} 30 to generate UAS-Ras1CA. Primers used here and hereafter are listed in Supplementary information, Table S1.
The DNA vector (Fil-GAL4 or UAS-Ras1CA) and helper plasmid pHA3PIG were injected into silkworm eggs at a concentration of 0.4 μg/μl for each plasmid 31. Positive G1 embryos were selected by DsRed fluorescence for Fil-GAL4 and EGFP fluorescence for UAS-Ras1CA under an Olympus MVX10 fluorescence stereomicroscope 30, 31. To confirm the positive hits detected by fluorescence microscopy, the insertion sites of the transgenic Fil-GAL4 and UAS-Ras1CA lines were also determined by inverse PCR 32. Multiple independent transgenic lines were used for Fil-GAL4 and UAS-Ras1CA. UAS-Ras1CA was crossed with Fil-GAL4 to generate the Fil-GAL4/UAS-Ras1CA lines, in which Ras1CA was specifically overexpressed in the posterior silk gland. The progeny of this cross showed four different phenotypes in terms of eye color (Figure 1): (1) neither DsRed2-positive nor EGFP-positive, wild type, [D(−)E(−)]; (2) only DsRed2-positive, GAL4 lines, [D(+)E(−)]; (3) only EGFP-positive, UAS lines, [D(−)E(+)]; and (4) both DsRed2-positive and EGFP-positive, GAL4/UAS lines, [D(+)E(+)].
Determination of mRNA levels and total DNA content
Total RNA was extracted using Trizol (Invitrogen) from the posterior or middle silk gland from the above four different phenotypes: [D(−)E(−)], [D(+)E(−)], [D(−)E(+)], [D(+)E(+)]. qPCR was used to determine mRNA levels as previously described 16. Total genomic DNA was extracted from the posterior silk gland at the early wandering stage using standard DNA SDS lysis-phenol extraction treatment after incubation with proteinase K. The DNA was further treated with RNAase and purified 32. Total DNA content was measured at OD 260 nm. Except where otherwise specified, all posterior silk gland samples were collected at the early wandering stage, when silkworms just begin to spin.
Ras1 activation assay and western blotting
Ras1 activity was assessed with a RAS activation assay kit purchased from Upstate Biotechnology. Western blotting was performed using standard methods as previously described 33. The primary antibodies used in this study included phospho-MAPK (Thr180/Tyr182, #9211, Cell Signaling), phosphor-InR (Tyr1150/1151, #3024, Cell Signaling), phospho-Akt (Ser505, #4054, Cell Signaling), phospho-S6K (Thr412, #07-018SP, Millipore), phospho-4EBP (Thr37/46, #9459, Cell Signaling), S6 (#2317, Cell Signaling), and α-tubulin (#AT819, Beyotime).
BrdU labeling and microscopy
BrdU labeling in the posterior silk gland was performed as previously described 34 with slight modification. In addition, 100 μg BrdU was injected into each larva on day 6 of the 5th instar. Those silkworms, which began to wander 24 h after BrdU injection, were selected for later staining experiments. Section samples were fixed in 5% formaldehyde for 45 min. After the DNA was denatured, the glands were incubated with anti-BrdU primary antibody (1:100; BD Biosciences) for 2 h and then with FITC-coupled goat anti-mouse secondary antibody (1:200; Jackson ImmuoResearch) for 2 h. BrdU labeled samples were evaluated under an Olympus Fluoview FV1000 confocal microscope.
Section samples for optical microcopy were observed using an Olympus IX71 inverted fluorescence microscope, and those for transmission electron microscopy were observed under a Hitachi HU-12 electron microscope at an accelerating voltage of 100 kV.
Statistics
Experimental data were statistically analyzed using ANOVA and Student's t-test using an SAS program.
References
Ribaut JM, de Vicente MC, Delannay X . Molecular breeding in developing countries: challenges and perspectives. Curr Opin Plant Biol 2010; 13:1–6.
Yoshiaki M, Hirovvo I, Kiyoshi S, Keiko I, Yoichi M, Shuji H . Studies on the protein synthesis in silkglands V. The relation of ribosomes to endoplasmic reticulum during fibroin synthesis. J Biochemi 1964; 55:623–628.
Tashiro Y, Morimoto T, Matsuura S, Nagata S . Studies on the posterior silk gland of the silkworm Bombyx mori I. Growth of posterior silk gland cells and biosynthesis of fibroin during the fifth larval instar. J Cell Biol 1968; 38:574–588.
Shigematsu H, Kurata K, Takeshita, H . Nucleic acids accumulation of silk gland of Bombyx mori in relation to silk protein. Comp Biochem Physiol B 1978; 61:237–242.
Blaes N, Couble P, Prudhomme JC . The programming of silk-gland development in Bombyx mori. I. Effects of experimental starvation on growth, silk production, and autolysis during the fifth larval instar studied by electron microscopy. Cell Tissue Res 1980; 213:311–324.
Akai HII . The structure and ultrastructure of the silk gland. Experientia 1983; 39:443–449.
Karnoub AE, Weinberg RA . Ras oncogenes: split personalities. Nat Rev Mol Cell Biol 2008; 9:517–531.
Caldwell, P.E ., Walkiewicz, M ., Stern, M . Ras activity in the Drosophila prothoracic gland regulates body size and developmental rate via ecdysone release. Curr Biol 2005; 15:1785–1795.
Ogura T, Tan A, Tsubota T, Nakakura T, Shiotsuki T . Identification and expression analysis of ras gene in silkworm, Bombyx mori. PLoS One 2009; 4:e8030.
Moriya K, Tsubota T, Ishibashi N, et al. Bombyx mori Ras proteins BmRas1, BmRas2 and BmRas3 are neither farnesylated nor palmitoylated but are geranylgeranylated. Insect Mol Biol 2010; 19:291–301.
Imamura M, Nakai J, Inoue S, Quan GX, Kanda T, Tamura T . Targeted gene expression using the GAL4/UAS system in the silkworm Bombyx mori. Genetics 2003; 165:1329–1340.
Tan AJ, Tanaka H, Tamura T, Shiotsuki T . Precocious metamorphosis in transgenic silkworms overexpressing juvenile hormone esterase. Proc Natl Acad Sci USA 2005; 102:11751–11756.
Oldham S, Hafen E . Insulin/IGF and target of rapamycin signaling: a TOR de force in growth control. Trends Cell Biol 2003; 13:79–85.
Tamura T, Thibert C, Royer C, et al. Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat Biotechnol 2000; 18: 81–84.
Duffy JB . GAL4 system in Drosophila: a fly geneticist's Swiss army knife. Genesis 2002; 34:1–15.
Liu Y, Zhou S, Ma L, et al. Transcriptional regulation of the insulin signaling pathway genes by starvation and 20-hydroxyecdysone in the Bombyx fat body. J Insect Physiol 2010; 56:1436–1444.
Xia Q, Guo Y, Zhang Z, et al. Complete resequencing of 40 genomes reveals domestication events and genes in silkworm (Bombyx). Science 2009; 326:433–436.
Xia Q, Cheng D, Duan J, et al. Microarray-based gene expression profiles in multiple tissues of the domesticated silkworm, Bombyx mori. Genome Biol 2007; 8:R162.
Edgar BA, Orr-Weaver TL . Endoreplication cell cycles: more for less. Cell 2001; 105:297–306.
Chang F, Steelman LS, Lee JT, et al. Signal transduction mediated by the Ras/Raf/MEK/ERK pathway from cytokine receptors to transcription factors: potential targeting for therapeutic intervention. Leukemia 2003; 17:1263–1293.
Adhikary S, Eilers M . Transcriptional regulation and transformation by Myc proteins. Nat Rev Mol Cell Biol 2005; 6:635–645.
Britton JS, Edgar BA . Environmental control of the cell cycle in Drosophila: nutrition activates mitotic and endoreplicative cells by distinct mechanisms. Development 1998; 125:2149–2158.
Frei C, Edgar BA . Drosophila cyclin D/Cdk4 requires Hif-1 prolyl hydroxylase to drive cell growth. Dev Cell 2004; 6:241–251.
Brogiolo W, Stocker H, Ikeya T, Rintelen F, Fernandez R, Hafen E . An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr Biol 2001; 11:213–221.
Wullschleger S, Loewith R, Hall MN . TOR signaling in growth and metabolism. Cell 2006; 124:471–484.
Bhaskar PT, Hay N . The two TORCs and Akt. Dev Cell 2007; 12:487–502.
Zhou S, Zhou Q, Liu Y, et al. Two Tor genes in the silkworm, Bombyx mori. Insect Mo Biol 2010; 19:727–735.
Kikuchi Y, Mori K, Suzuki S, Yamaguchi K, Mizuno S . Structure of the Bombyx mori fibroin light-chain-encoding gene: upstream sequence elements common to the light and heavy chain. Gene 1992; 110:151–158.
Brand AH, Perrimon N . Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993; 118:401–415.
Dai H, Ma L, Wang J, Jiang RJ, Wang Z, Fei J . Knockdown of ecdysis-triggering hormone gene with a binary UAS/GAL4 RNA interference system leads to lethal ecdysis deficiency in silkworm. Acta Biochim Biophys Sin 2008; 40:790–795.
Zhao A, Zhao T, Zhang Y, et al. New and highly efficient expression systems for expressing selectively foreign protein in the silk glands of transgenic silkworm. Transgenic Res 2010; 19:29–44.
Hediger M, Niessen M, Wimmer EA, Dübendorfer A, Bopp D . Genetic transformation of the housefly Musca domestica with the lepidopteran derived transposon piggyBac. Insect Mol Biol 2001; 10:113–119.
Tian L, Guo E, Dian Y, et al. Genome wide regulation of innate immunity by juvenile hormone and 20-hydroxyecdysone in the Bombyx fat body. BMC Genomics 2010; 11:549.
Li S . Identification of iron-loaded ferritin as an essential mitogen for cell proliferation and postembryonic development in Drosophila. Cell Res 2010; 20:1148–1157.
Acknowledgements
The transgenic silkworm Fil-GAL4/UAS-Ras1CA has been protected by the Chinese Intellectual Property Deputy under the patent 201010118072.8. This study was supported by 2007AA10Z155, KSCX-YW-N-009, 30770271, 10JC1416700, 2007CB947100, 2006CB943902, 30870299, 2006AA10A119, and Hundred Talent Project to SL as well as 2005CB121000 to QX. We are grateful to Drs Jian Fei, Zhugang Wang, and Hongjiu Dai in Shanghai Research Center for Model Organisms for their support of the initial transgenic silkworm studies. We thank Dr David Stanley for improving this manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
( Supplementary information is linked to the online version of the paper on the Cell Research website.)
Supplementary information
Supplementary information, Figure S1
MAPK phosphorylation level correlates and slightly precedes the growth rate of posterior silk gland. (PDF 64 kb)
Supplementary information, Figure S2
Inhibition of PI3K110 activity reduces silk gland size. (PDF 428 kb)
Supplementary information, Figure S3
Increasing posterior silk gland size by Ras1CA overexpression greatly improves silk yield. (PDF 108 kb)
Supplementary information, Figure S4
BrdU labeling in the posterior silk gland from [D(+)E(+)] (A') is stronger than those in the controls [D(−)E(−)] (A). (PDF 76 kb)
Supplementary information, Figure S5
Ras1 activation increases the posterior silk gland size observed under optical microscopy. (PDF 22 kb)
Supplementary information, Figure S6
A proposed model shows that increasing posterior silk gland size and stimulating fibroin protein synthesis by Ras1CA overexpression in the transgenic silkworm Fil-GAL4/UAS-Ras1CA. (PDF 91 kb)
Supplementary information, Table S1
All primers used in this paper (PDF 7 kb)
Rights and permissions
About this article
Cite this article
Ma, L., Xu, H., Zhu, J. et al. Ras1CA overexpression in the posterior silk gland improves silk yield. Cell Res 21, 934–943 (2011). https://doi.org/10.1038/cr.2011.36
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cr.2011.36
Keywords
This article is cited by
-
Performance of some newly produced multivoltine strains of Bombyx mori (Linnaeus) for commercially vibrant qualitative and quantitative traits
International Journal of Tropical Insect Science (2021)
-
A transcriptome atlas of silkworm silk glands revealed by PacBio single-molecule long-read sequencing
Molecular Genetics and Genomics (2020)
-
Insights into the regulatory characteristics of silkworm fibroin gene promoters using a modified Gal4/UAS system
Transgenic Research (2019)
-
New insight into the mechanism underlying the silk gland biological process by knocking out fibroin heavy chain in the silkworm
BMC Genomics (2018)
-
Extraordinary Mechanical Properties of Composite Silk Through Hereditable Transgenic Silkworm Expressing Recombinant Major Ampullate Spidroin
Scientific Reports (2018)
