
Abstract
Vesicular stomatitis virus (VSV) is neuropathogenic in rodents but can be attenuated 50-fold by engineering the mouse interferon-beta (IFN-β) gene into its genome. Intravenously administered VSVs encoding IFN-β have potent activity against subcutaneous tumors in the 5TGM1 mouse myeloma model, without attendant neurotoxicity. However, when 5TGM1 tumor cells were seeded intravenously, virus-treated mice with advanced myeloma developed clinical signs suggestive of meningoencephalitis. Co-administration of a known active antimyeloma agent did not prolong survival, further suggesting that deaths were due to viral toxicity, not tumor burden. Histological analysis revealed that systemically administered 5TGM1 cells seed to the CNS, forming meningeal tumor deposits, and that VSV infects and destroys these tumors. Death is presumably a consequence of meningeal damage and/or direct transmission of virus to adjacent neural tissue. In light of these studies, extreme caution is warranted in clinical testing of attenuated VSVs, particularly in patients with CNS tumor deposits.
Similar content being viewed by others
Introduction
Oncolytic viruses selectively target tumor cells by exploiting the differences between tumor and normal cells.1 A number oncolytic viruses, including reovirus, mumps, adenovirus, measles virus, herpes simplex virus, poliovirus and vaccinia virus have entered clinical trials for use as anticancer agents.2, 3 Additionally, oncolytic vesicular stomatitis viruses (VSVs) have shown great potential for the treatment of a variety of tumors, including glioblastoma, sarcoma, colon carcinoma, ovarian carcinoma, B-cell lymphoma and multiple myeloma,4, 5 and a human clinical trial in patients with hepatocellular carcinoma is now underway (ClinicalTrials.gov identifier: NCT01628640).
VSV is a bullet-shaped, negative-sense, single-stranded RNA virus of the Rhabdoviridae family that does not integrate its genome into the host cell.4 The genome of VSV codes for five proteins, namely the nucleocapsid (N), the phosphoprotein (P), the peripheral matrix protein (M), the surface glycoprotein (G) and the large protein or polymerase (L).6 This virus, which is typically a pathogen of livestock and relatively nonpathogenic to humans, can replicate to high titers in a wide variety of cell types, including tumor cells.7, 8, 9
Although VSV has shown great potential as a potent oncolytic, this virus is also known to be neurotoxic. Following intranasal injection, for example, VSV infects olfactory neurons, which subsequently leads to infection of the olfactory bulb and the central nervous system (CNS), causing lethal encephalitis in mice.10, 11 Encephalitis in mice has also been reported following intraperitoneal, intranasal, intramuscular, subcutaneous and intravenous injection of VSV.12 VSV-induced neurotoxicity has been shown to cause lethal encephalitis in mice, hamsters and non-human primates,8, 13, 14, 15, 16, 17 with lethal encephalitis in mice typically occurring within 10 days of infection with VSV.11
The neurotoxic effects of VSV can be inhibited by viral mutations or by insertion of neuroattenuating genes into the genome of this virus. For example, VSV neuroattenuation has been achieved by repositioning the M cistron, G truncations or inserting a picornaviral internal ribosomal entry site to attenuate M protein expression,18, 19, 20 by viral expression of p53 and of various cytokines21, 22, 23 or by direct mutation of the M protein sequence (VSVΔM51).24, 25 VSVΔM51 is attenuated in normal, interferon (IFN)-responsive cells but maintains oncolytic activity in tumor cells defective of IFN signaling.25 Along these same lines, the interferon (IFN)-β gene has been introduced into the VSV genome. The production of IFN-β following infection with virus leads ultimately to inhibition of viral replication.26 IFN pathways are commonly defective in tumor cells, however, rendering these cells resistant to the antiviral effects induced by IFN-β expression.4 Thus, VSV expressing IFN-β is attenuated in non-malignant cells while retaining its oncolytic activity, and the virally encoded IFN-β has also been shown to enhance the therapeutic efficacy of VSV treatment.27, 28, 29, 30, 31 Unfortunately, however, these neuroattenuated viruses can still be lethal at high titers.32
In this report, we studied a neuroattenuated VSV in a systemic myeloma model. We tested the safety and efficacy of a recombinant VSV coding for both murine IFN-β and the sodium iodide symporter (NIS; VSV-mIFNβ-NIS). This virus, which showed therapeutic benefits both in subcutaneous and early-stage systemic mouse myeloma models, did not prolong survival of mice with advanced systemic myeloma. The data presented here show that even an attenuated VSV can cause lethal meningoencephalitis when CNS tumor deposits are present.
Materials and methods
In vivo experiments
Animal protocols were approved by the Mayo Clinic Institutional Care and Use Committee. Female C57BL/KaLwRijHsd mice were obtained from Harlan Laboratories (Netherlands), and syngeneic murine myeloma 5TGM1 cells were implanted.33 For subcutaneous studies, 5 × 106 5TGM1 cells were implanted, and VSV (108 TCID50) was administered intravenously (IV; via the tail vein) 14 days later. Systemic myeloma was established by systemic (IV, tail vein) injection of 107 5TGM1 cells. Twenty-eight days post systemic 5TGM1 administration, mice were treated with VSV (108 TCID50; IV). Mice were given cyclophosphamide (100 mg kg−1; intraperitoneal injection) on day 30 post 5TGM1 administration.
Viruses
Mice were treated with one of two recombinant VSVs. The VSV-Luc plasmid, provided by Glen Barber from the University of Miami, contains the firefly luciferase reporter (inserted between the G and L genes of VSV). Plasmid DNA for VSV-mIFNβ-NIS was generated in our laboratory and encodes the mouse interferon-beta gene (inserted between M and G) and the sodium iodide symporter (NIS; inserted between G and L). The recombinant viruses were propagated as previously described.34
Histology and immunofluorescence
Tumors, brains and spinal cords collected from mice were placed in 10% neutral-buffered formalin and paraffin embedded. Serial sections were either stained with hematoxylin and eosin (H&E) or used for VSV detection via immunofluorescent techniques. Briefly, deparaffinization and rehydration were performed by placing slides in three changes of xylene followed by washes in graded alcohol. Slides were then washed in PBS, and antigen retrieval was performed using 1X Target Retrieval Solution (Dako, Carpinteria, CA, USA) according to the manufacturer’s protocol. Following antigen retrieval, samples were incubated in block solution (5% normal goat serum in 0.2% Triton-X/PBS) for 20 min. Next, a rabbit antibody against VSV was added, and the slides were subsequently washed with PBS. Samples were then incubated with an Alexa Fluor 555 goat anti-rabbit secondary antibody (Invitrogen, Carlsbad, CA, USA) and counterstained with Hoechst. Fluorescent images were obtained with a Zeiss Axiovert microscope.
Luciferase detection
Luciferase activity was analyzed in mice 24 h after treatment with VSV-Luc. Mice were anesthetized by isoflurane inhalation following intraperitoneal injection with D-luciferin (3 mg per mouse; Gold Biotechnology). Approximately 10 min after luciferin injection, mice were placed on the imaging platform and analysis was performed using a Xenogen Living Systems 2.60.2 software.
Statistical analysis
Survival data were analyzed using GraphPad Prism (v. 5.0a) software (GraphPad Software, San Diego, CA, USA). Kaplan–Meier survival curves were plotted, and statistical differences were determined by log-rank (Mantel-Cox) analysis.
Results
Vesicular stomatitis virus effectively targets tumors but does not enhance survival of mice with systemic myeloma
Pre-clinically, vesicular stomatitis virus (VSV) has shown great promise as an oncolytic virus.4, 5 Initially, to test the oncolytic efficacy of VSV in myeloma, we implanted 5TGM1 murine myeloma cells (5 × 106 cells subcutaneously) into syngeneic, immunocompetent C57BL/KaLwRij mice. Fourteen days post implantation, mice were treated with PBS (control) or with VSV-mIFNβ-NIS, a neuroattenuated VSV encoding mouse interferon (mIFN)-beta and the sodium iodide symporter (NIS). As shown in Figure 1a, mice treated with VSV-mIFNβ-NIS survived significantly longer than control-treated mice (P=0.0084).

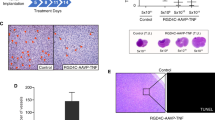
Treatment with vesicular stomatitis virus (VSV) effectively targets tumor cells but does not enhance the survival of mice with orthotopic myeloma. (a) 5TGM1 myeloma cells were implanted subcutaneously into C57BL/KaLwRij mice. Mice were treated with VSV-mIFN-NIS (IV, 108 TCID50) at day 14. VSV-treated mice (n=10) showed a statistically significant increase (P=0.0084) in survival when compared with control-treated mice (n=7). (b) 5TGM1 cells were injected systemically (IV, tail vein) into C57BL/KaLwRij mice. Mice were treated with VSV-mIFN-NIS (IV, 108 TCID50) at day 28. No difference in survival (P=0.332) was seen between control-treated (n=5) and VSV-treated mice (n=5). (c) H&E analysis of palpable tumors obtained from mice with advanced systemic myeloma. The PBS-treated (control) tumor contains mostly viable tumor cells, whereas the tumor from the VSV-treated mouse consists mainly of necrotic cells. (d) Immunofluorescent analysis of the same tumors shown in panel (c). Staining for VSV is in red; Hoechst is shown in blue.
As myeloma is a disseminated malignancy, we next moved to a systemic myeloma model. C57BL/KaLwRij mice were given an intravenous injection (via the tail vein) of 107 5TGM1 myeloma tumor cells, and 28 days later were treated with VSV-mIFNβ-NIS or PBS (control). In contrast to the results seen in mice with subcutaneous tumors, there was no difference in survival between control-treated mice and mice treated with VSV-mIFNβ-NIS (Figure 1b).
Mice with advanced systemic myeloma occasionally develop palpable abdominal and pelvic tumors. We collected and analyzed tumors from control- and VSV-treated mice. As shown in Figure 1c, hematoxylin and eosin (H&E) staining shows mostly viable tumor cells in the tumor from the PBS-treated mouse. Conversely, the solid tumor obtained from the mouse treated with VSV consists mainly of necrotic cells (Figure 1c). In addition, using immunofluorescent analysis, VSV was detected in this tumor (Figure 1d; VSV staining is shown in red and Hoechst staining in blue). In total, these results show that, although VSV can efficiently target myeloma tumor cells, this virus does not prolong survival of mice with advanced, orthotopic myeloma.
VSV-treated mice become moribund for different reasons than control-treated mice
We observed that treatment with oncolytic VSV enhanced the survival of mice with subcutaneous myeloma tumors, but did not provide a significant survival advantage to mice with advanced systemic myeloma (Figures 1a and b). A separate set of mice with orthotopic myeloma was treated with VSV containing a luciferase reporter (VSV-Luc) or PBS. The majority of the control-treated mice (86%) died or were euthanized because of tumor progression leading to cord compression and the onset of hind-limb paralysis, which inhibited their ability to access food and water (Table 1). On the other hand, the majority of VSV-treated mice (56%) were euthanized because of withdrawn, lethargic behavior (Table 1). These mice had labored breathing and withdrew to a corner of their cage, did not eat or drink, and thus lost weight rapidly.
On the basis of the observed differences in cause of death (Table 1) and the results showing that VSV can efficiently target peripheral tumors in these mice (Figures 1c and d), we hypothesized that VSV-treated mice with advanced, orthotopic myeloma were not dying of tumor burden, but rather of VSV-induced neurotoxicity.
A therapeutically effective dose of cyclophosphamide does not prolong survival of VSV-treated mice
Previously, it was reported that cyclophosphamide (CPA) treatment enhances the survival of mice with advanced myeloma.35 We hypothesized that VSV is not efficacious in the present model because of the neurotoxic effects of this virus; therefore, an effective dose of CPA will not be able to prolong the survival of VSV-treated mice because they are dying of neurotoxicity and not tumor burden. Tumor-bearing mice were separated into four treatment groups: control (PBS); VSV alone; CPA alone; and VSV+CPA. VSV was given 28 days after 5TGM1 injection, and a single dose of CPA was administered 48 h post VSV treatment (Day 30 post-5TGM1). As shown in Figure 2, mice treated with CPA alone had a significant survival advantage compared to mice treated with the combination of VSV+CPA (P=0.0143). As a therapeutically effective dose of CPA was not able to rescue VSV-treated mice, these results suggest that the mice are not dying because of tumor burden but rather because of the adverse effects of VSV treatment.

Cyclophosphamide treatment does not prolong survival of VSV-treated mice. 5TGM1 cells were injected systemically (IV, tail vein) into C57BL/KaLwRij mice. Mice (n=3 per group) were treated with PBS or VSV-mIFN-NIS (IV, 108 TCID50) at 28 days post-5TGM1 injection; 2 days later, mice were then treated with PBS or cyclophosphamide (CPA; 100 mg kg−1). A survival curve comparing mice treated with PBS, VSV, CPA or VSV+CPA is shown. CPA-treated mice have a significant survival advantage over mice in all other treatment groups.
Tumor cells and VSV are detected in the meninges of mice with advanced systemic myeloma
We decided to further analyze the tissues of the central nervous system (CNS) of untreated and VSV-treated mice to look for evidence of VSV-induced neurotoxicity. Therefore, at the time of death, we collected the brains and spinal cords of experimental animals and performed histopathologic analysis. Interestingly, tumor cells were observed in the meninges of many of these mice. Specifically, 86% (6/7) of the brains of tumor-bearing mice not exposed to VSV contained myeloma cells in the meninges (Figure 3a). However, tumor cells were only found in 20% (2/10) of the meninges surrounding the brains of mice treated with VSV (Figure 3a). Further, although viable tumor cells were detected in the meninges of control mice, the myeloma cells detected in the meninges of the VSV-treated mice were necrotic (Figure 3b). We hypothesized that these tumor cells were necrotic because of the oncolytic activity of VSV and, using immunofluorescent techniques, we were also able to detect VSV in the meninges (Figure 3c; staining for VSV is shown in red and Hoechst staining in blue). Additionally, inflammatory cells, including lymphocytes and macrophages, were found infiltrating the spinal cord tissue of a VSV-treated mouse, but none were detected in the spinal cord of a control-treated mouse (Figure 3d).

Tumor cells are detected in the meninges of mice with advanced systemic myeloma. (a) Bar graph comparing the percentage of VSV-treated (n=10) versus non-VSV-treated (n=7) mice with 5TGM1 tumor cells detected in the meninges of the brain. Tumor cells detected in VSV-treated mice were necrotic. (b) H&E analysis of meningeal myeloma tumors obtained from mice with systemic advanced myeloma. The PBS-treated (control) tumors contain mostly viable tumor cells, whereas the tumors from the mice treated with VSV consist of mainly necrotic cells. (c) Immunofluorescent analysis of the same tumors shown in panel (b). Staining for VSV is in red; Hoechst is shown in blue. (d) H&E analysis of the spinal cord of a control- and a VSV-treated mouse. Arrows point to representative inflammatory cells.
Luciferase activity is detected in the tumors and CNS of mice treated with VSV-Luc
To further confirm that VSV is targeting the tumor cells and also spreads to the CNS, we analyzed the distribution of VSV in mice using a VSV encoding luciferase (VSV-Luc) and non-invasive imaging techniques. VSV-Luc was administered systemically to tumor-bearing mice (that is, mice with advanced systemic myeloma) as well as to mice without 5TGM1 tumors (control). Twenty-four hours after VSV-Luc treatment, mice were injected with D-luciferin, and luciferase activity was analyzed (Figure 4). Luciferase activity was detected only at the site of injection (tail vein) in the control mouse. In comparison, luciferase expression in tumor-bearing mice was visible at the site of injection as well as at the sites of peripheral tumors, indicating that VSV does indeed target tumor cells. Further, luciferase activity was also observed in the spinal cord and brains of these mice, providing further evidence of VSV-induced neurotoxicity. In total, our data indicate that orthotopic 5TGM1 tumors, which seed to the CNS to form meningeal tumor deposits, are infected and rapidly destroyed by VSV, leading to fatal neurotoxicity.

Luciferase is detected in the tumors and CNS of tumor-bearing mice treated with VSV-Luc. PBS (Control) or 5TGM1 myeloma cells were injected systemically (IV, tail vein) into C57BL/KaLwRij mice (n=3 per group). Mice were treated with VSV-Luc (IV, tail vein; 108 TCID50) at day 28. VSV-Luc is detected in the tumors as well as the CNS of tumor-bearing mice. Arrows are pointing to peripheral tumors that developed in these mice.
Discussion
In this report, we have shown that VSV-mIFNβ-NIS treatment enhances the survival of mice with subcutaneous, syngeneic myeloma tumors. This virus, however, did not prolong survival of mice with advanced, orthotopic myeloma, even though it effectively targeted peripheral tumors. Further, a therapeutically efficacious dose of cyclophosphamide did not prolong the survival of VSV-treated mice, indicating that these mice are dying of the adverse effects of VSV and not of tumor burden. Histological analysis of CNS tissues showed myeloma cells in the meninges of mice with systemic myeloma. However, only necrotic tumor cells were detected in the meninges of VSV-treated mice, and VSV was also detected in the meninges via immunofluorescent analysis. Further, we detected inflammatory cells in the spinal cord tissue of a VSV-treated mouse but not of a control-treated mouse. In total, these results show that systemically administered VSV infects and destroys 5TGM1 cells that have seeded to the CNS and may then spread from the tumor to adjacent neural tissue, ultimately leading to fatal neurotoxicity.
The differences seen in the cause of death in VSV- and control-treated mice, in combination with the results of the cyclophosphamide rescue experiment, suggest that VSV-treated mice are indeed dying of neurotoxicity and not of tumor burden. In further support of this conclusion, very early reports analyzing VSV-induced neurotoxicity showed that nasal instillation of VSV into 15-day-old Rockefeller Institute albino mice induced lethal encephalitis within 10 days,11 and VSV (0.2ml of a 1:200 dilution of virus collected from fresh tissue in broth) injected intravenously via the tail vein caused paralysis within 5–6 days and death 24 h later.12 Similar to these reports, as seen in Figure 1b, Table 1 and Figure 2, C57BL/KaLwRij mice systemically infected with VSV (via IV, tail vein injection) all were euthanized within 10 days of VSV treatment. The majority of the mice were euthanized because of lethargic, withdrawn behavior, suggestive of encephalitis.
The combination of necrotic tumor cells and VSV detected in the meninges (Figures 3b and c), and inflammatory cells infiltrating the blood vessels and tissue of the spinal cord (Figure 3d), suggest that VSV is efficiently targeting and killing tumor cells within the meninges and may then be spreading directly to adjacent CNS tissue and causing fatal meningoencephalitis. In the absence of direct proof that VSV is spreading to the CNS, it remains possible, however, that meningoencephalitis is not the primary cause of death in these mice. For example, VSV may be causing tumor lysis syndrome, whereby tumor cells are killed rapidly, causing large amounts of intracellular components to begin to circulate systemically.36 Rapid tumor lysis can be fatal and might be a contributory factor in this model because tumor destruction does occur very rapidly, although detailed analyses of the blood of VSV-treated mice with 5TGM1 tumors do not support this hypothesis (data not shown). The destruction of tumor cells adjacent to the CNS, loss of meningeal integrity, or meningeal inflammation due to tumor cell deposits within that area, might also explain the demise of these mice. These causes of death are considered less likely, however, because the survival of mice with advanced systemic myeloma (that is, mice with tumor cell deposits in the meninges) could be prolonged with a single dose of cyclophosphamide (Figure 2).
Here we have shown that VSV induces neurotoxicity in the advanced systemic myeloma model. The neurotoxic effects may be model specific because of the presence of tumor cells in the meninges. Luciferase activity was not detected in the brains of non-tumor-bearing mice given the same dose of VSV-Luciferase (Figure 4). Further, tumor cells were not detected in the meninges of mice with subcutaneous tumors, and neurotoxicity was not evident in these mice, although they were treated with the same dose (108 TCID50) of virus. Additionally, treating mice with VSV at an earlier time (21 days versus 28 days) significantly prolonged survival, and tumor cells were not detected in the meninges of these mice.30
Model specificity of neurotoxicity caused by VSV exposure has also been demonstrated elsewhere. In a report by Olitsky et al.,12 young (15-day-old) Rockefeller Institute albino mice developed encephalitis following intraperitoneal, intranasal, intramuscular, subcutaneous and intravenous VSV injection. Conversely, 1-year-old mice of the same strain were relatively resistant to the neurotoxic effects of VSV, although these mice had no prior exposure to this virus.12 This same group also observed that intranasally injected VSV does not induce myelitis or encephalitis in guinea pigs, regardless of age.37 As another example, cynomolgus macaques inoculated intranasally with VSV (107 PFU) showed no signs of neuroinvasion, but intrathalamic injection of the same dose of VSV caused neurotoxicity resulting in declining health and euthanasia at 6-7 days following administration of the virus.8 Finally, although neurotoxicity is seen experimentally, natural infection of livestock is non-lethal and VSV infection in humans is typically asymptomatic.
Neuroattenuation of VSV is the key to successfully moving this virus into the clinic for use as an anticancer agent. VSV neuroattenuation has been achieved by mutating the M proteins of this virus. In 2003, Stojdl et al.25 found that VSV containing a single amino-acid substitution (M51R) was attenuated approximately 80-fold compared with wild-type VSV. Importantly, the oncolytic activity of this mutant virus was still maintained because of defective interferon (IFN) signaling in tumor cells.25 This group also found that a deletion of methionine 51 (VSVΔM51) similarly reduced neurotoxicity, and later studies using this neuroattenuated virus have shown that it can safely and effectively treat mice with multifocal and invasive gliomas, subcutaneous myeloma tumors and metastatic breast cancer.24, 25, 38, 39 Similarly, attenuated neurotoxicity has also been demonstrated when the IFN-β gene is directly engineered into the genome of VSV (VSV-IFNβ). Intravenous injection of VSV-IFNβ into BALB/c mice, for example, determined that this virus was 50 times more attenuated than wild-type VSV because of IFN-β expression.27 Also, VSV-(mouse)IFNβ injected intratumorally into SCID mice bearing mesothelioma tumors attenuated neurotoxicity and enhanced therapeutic efficacy when compared with mice treated with VSV-(human)IFNβ,29 and rhesus macaques injected intrahepatically with VSV-IFNβ did not show any adverse neurologic events with doses up to 1010 TCID50.32
Although neuroattenuation of VSV is promising, even attenuated viruses can be lethal if sufficient virus gains access to the CNS. In this report, we show that intravenous administration of VSV-mIFNβ is lethal to C57BL/KaLwRij mice with advanced systemic myeloma. Further, Jenks et al.32 reported that no adverse events were observed in Buffalo rats treated with 1010 TCID50 of VSV-IFNβ, but neurotoxicity was observed in 2 out of 12 BALB/c mice injected intrahepatically with this same virus at a dose of 2.95 × 109 TCID50. Along these same lines, a high titer of VSV-IFNβ (108 PFU) injected intravenously induced lethal neurotoxicity in two out of five BALB/c mice.27 Thus, caution is warranted even when the viruses tested are neuroattenuated.
The neuroattenuated VSVΔM51 has been found to prolong the survival of nude mice bearing U87 metastatic gliomas, with no evidence of neurotoxicity.24 Interestingly, in this report, minor VSV antigen staining was also detected in the meninges of mice 24 and 72 h after intravenous injection (5 × 108 PFU) of this virus, but no neurotoxicity was reported. The differences in neurotoxicity observed in this model and the advanced systemic myeloma model could be due to differences in the location of the tumor and the ability of the virus to spread following tumor cell lysis. Lun et al.24 report that GFP-labeled VSVΔM51 does not spread to the surrounding normal brain tissue, but instead remains confined to the glioma tumor cells. On the other hand, in the myeloma model, the tumor cell deposits are located within the meninges of the CNS, and our data suggest that VSV infects and destroys these tumor cells, then spreads to and infects CNS tissue. The differences in neurotoxicity seen between the glioma and myeloma models could be due to differences in the degree of stimulation of the innate immune response following virus infection. It is possible that IFN production and innate immunity following VSV infection is greater in the glioma model compared with the myeloma model, leading to a greater inhibition of viral propagation in the gliomas, reducing the risk of viral spread to adjacent brain.
In conclusion, neurotoxicity caused by VSV treatment remains a concern when considering treating patients with this virus. Importantly, myeloma cells rarely seed to the CNS of myeloma patients, as was seen in the advanced systemic mouse model, suggesting that neurotoxicity leading to lethal encephalitis may not be a concern in the majority of this group of patients. However, many tumors do metastasize to the CNS, and extreme caution must be taken in the clinical testing of VSV, including neuroattenuated recombinant VSVs, in patients with CNS tumor deposits.
References
Parato KA, Senger D, Forsyth PA, Bell JC . Recent progress in the battle between oncolytic viruses and tumours. Nat Rev Cancer 2005; 5: 965–976.
Liu TC, Galanis E, Kirn D . Clinical trial results with oncolytic virotherapy: a century of promise, a decade of progress. Nat Clin Pract Oncol 2007; 4: 101–117.
Russell SJ, Peng KW, Bell JC . Oncolytic virotherapy. Nat Biotechnol 2012; 30: 658–670.
Barber GN . Vesicular stomatitis virus as an oncolytic vector. Viral Immunol 2004; 17: 516–527.
Lichty BD, Stojdl DF, Taylor RA, Miller L, Frenkel I, Atkins H et al. Vesicular stomatitis virus: a potential therapeutic virus for the treatment of hematologic malignancy. Hum Gene Ther 2004; 15: 821–831.
Assenberg R, Delmas O, Morin B, Graham SC, De Lamballerie X, Laubert C et al. Genomics and structure/function studies of Rhabdoviridae proteins involved in replication and transcription. Antiviral Res 2010; 87: 149–161.
Barber GN . VSV-tumor selective replication and protein translation. Oncogene 2005; 24: 7710–7719.
Johnson JE, Nasar F, Coleman JW, Price RE, Javadian A, Draper K et al. Neurovirulence properties of recombinant vesicular stomatitis virus vectors in non-human primates. Virology 2007; 360: 36–49.
Rodriguez LL . Emergence and re-emergence of vesicular stomatitis in the United States. Virus Res 2002; 85: 211–219.
Reiss CS, Plakhov IV, Komatsu T . Viral replication in olfactory receptor neurons and entry into the olfactory bulb and brain. Ann N Y Acad Sci 1998; 855: 751–761.
Sabin AB, Olitsky PK . Influence of host factors on neuroinvasiveness of vesicular stomatitis virus: I. effect of age on the invasion of the brain by virus instilled in the nose. J Exp Med 1937; 66: 15–34.
Olitsky PK, Sabin AB, Cox HR . An acquired resistance of growing animals to certain neurotropic viruses in the absence of humoral antibodies or previous exposure to infection. J Exp Med 1936; 64: 723–737.
Bi Z, Barna M, Komatsu T, Reiss CS . Vesicular stomatitis virus infection of the central nervous system activates both innate and acquired immunity. J Virol 1995; 69: 6466–6472.
Fournier JG, Robain O, Cerutti I, Tardivel I, Chany-Fournier F, Chany C . Detection of vesicular stomatitis virus (VSV) RNA in the central nervous system of infected mice by in situ hybridization. Acta Neuropathol 1988; 75: 554–556.
Fultz PN, Shadduck JA, Kang CY, Streilein JW . Genetic analysis of resistance to lethal infections of vesicular stomatitis virus in Syrian hamsters. Infect Immun 1981; 32: 1007–1013.
Huneycutt BS, Plakhov IV, Shusterman Z, Bartido SM, Huang A, Reiss CS et al. Distribution of vesicular stomatitis virus proteins in the brains of BALB/c mice following intranasal inoculation: an immunohistochemical analysis. Brain Res 1994; 635: 81–95.
Plakhov IV, Arlund EE, Aoki C, Reiss CS . The earliest events in vesicular stomatitis virus infection of the murine olfactory neuroepithelium and entry of the central nervous system. Virology 1995; 209: 257–262.
Ammayappan A, Nace R, Peng KW, Russell SJ . Neuroattenuation of vesicular stomatitis virus through picornaviral internal ribosome entry sites. J virol 2013; 87: 3217–3228.
Wollmann G, Rogulin V, Simon I, Rose JK, van den Pol AN . Some attenuated variants of vesicular stomatitis virus show enhanced oncolytic activity against human glioblastoma cells relative to normal brain cells. J virol 2010; 84: 1563–1573.
Cooper D, Wright KJ, Calderon PC, Guo M, Nasar F, Johnson JE et al. Attenuation of recombinant vesicular stomatitis virus-human immunodeficiency virus type 1 vaccine vectors by gene translocations and g gene truncation reduces neurovirulence and enhances immunogenicity in mice. J virol 2008; 82: 207–219.
Heiber JF, Barber GN . Vesicular stomatitis virus expressing tumor suppressor p53 is a highly attenuated, potent oncolytic agent. J virol 2011; 85: 10440–10450.
Ramsburg E, Publicover J, Buonocore L, Poholek A, Robek M, Palin A et al. A vesicular stomatitis virus recombinant expressing granulocyte-macrophage colony-stimulating factor induces enhanced T-cell responses and is highly attenuated for replication in animals. J virol 2005; 79: 15043–15053.
Miller J, Bidula SM, Jensen TM, Reiss CS . Cytokine-modified VSV is attenuated for neural pathology, but is both highly immunogenic and oncolytic. Int J Infereron Cytokine Mediator Res 2009; 1: 15–32.
Lun X, Senger DL, Alain T, Oprea A, Parato K, Stojdl D et al. Effects of intravenously administered recombinant vesicular stomatitis virus (VSV(deltaM51)) on multifocal and invasive gliomas. J Natl Cancer Inst 2006; 98: 1546–1557.
Stojdl DF, Lichty BD, tenOever BR, Paterson JM, Power AT, Knowles S et al. VSV strains with defects in their ability to shutdown innate immunity are potent systemic anti-cancer agents. Cancer Cell 2003; 4: 263–275.
Stark GR, Kerr IM, Williams BR, Silverman RH, Schreiber RD . How cells respond to interferons. Annu Rev Biochem 1998; 67: 227–264.
Obuchi M, Fernandez M, Barber GN . Development of recombinant vesicular stomatitis viruses that exploit defects in host defense to augment specific oncolytic activity. J Virol 2003; 77: 8843–8856.
Saloura V, Wang LC, Fridlender ZG, Sun J, Cheng G, Kapoor V et al. Evaluation of an attenuated vesicular stomatitis virus vector expressing interferon-beta for use in malignant pleural mesothelioma: heterogeneity in interferon responsiveness defines potential efficacy. Hum Gene Ther 2010; 21: 51–64.
Willmon CL, Saloura V, Fridlender ZG, Wongthida P, Diaz RM, Thompson J et al. Expression of IFN-beta enhances both efficacy and safety of oncolytic vesicular stomatitis virus for therapy of mesothelioma. Cancer Res 2009; 69: 7713–7720.
Naik S, Nace R, Barber GN, Russell SJ . Potent systemic therapy of multiple myeloma utilizing oncolytic vesicular stomatitis virus coding for interferon-beta. Cancer gene therapy 2012; 19: 443–450.
Naik S, Nace R, Federspiel MJ, Barber GN, Peng KW, Russell SJ . Curative one-shot systemic virotherapy in murine myeloma. Leukemia 2012; 26: 1870–1878.
Jenks N, Myers R, Greiner SM, Thompson J, Mader EK, Greenslade A et al. Safety studies on intrahepatic or intratumoral injection of oncolytic vesicular stomatitis virus expressing interferon-beta in rodents and nonhuman primates. Hum Gene Ther 2010; 21: 451–462.
Oyajobi BO, Franchin G, Williams PJ, Pulkrabek D, Gupta A, Munoz S et al. Dual effects of macrophage inflammatory protein-1alpha on osteolysis and tumor burden in the murine 5TGM1 model of myeloma bone disease. Blood 2003; 102: 311–319.
Kelly EJ, Nace R, Barber GN, Russell SJ . Attenuation of vesicular stomatitis virus encephalitis through microRNA targeting. J Virol 2010; 84: 1550–1562.
Sharabi A, Fau—Laronne-Bar-On A, Laronne-Bar-On A, Fau—Meshorer A, Meshorer A, Fau—Haran-Ghera N et al. Chemoimmunotherapy reduces the progression of multiple myeloma in a mouse model. (1940-6215 (Electronic)).
Muslimani A, Chisti MM, Wills S, Nadeau L, Zakalik D, Daw H et al. How we treat tumor lysis syndrome. Oncology 2011; 25: 369–375.
Sabin AB, Olitsky PK. . Influence of host factors on neuroinvasiveness of vesicular stomatitis virus: Iv. Variations in neuroinvasiveness in different species. J Exp Med 1938; 67: 229–249.
Ebert O, Harbaran S, Shinozaki K, Woo SL . Systemic therapy of experimental breast cancer metastases by mutant vesicular stomatitis virus in immune-competent mice. Cancer Gene Ther 2005; 12: 350–358.
Goel A, Carlson SK, Classic KL, Greiner S, Naik S, Power AT et al. Radioiodide imaging and radiovirotherapy of multiple myeloma using VSV(Delta51)-NIS, an attenuated vesicular stomatitis virus encoding the sodium iodide symporter gene. Blood 2007; 110: 2342–2350.
Acknowledgements
We thank Dr Ronald Marler and his laboratory for assisting with the IHC slide preparation and analysis.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
SN, KWP, MJF and SJR and Mayo Clinic own equity and have financial interests in Omnis Pharma, an oncolytic VSV company.
Rights and permissions
About this article
Cite this article
Yarde, D., Naik, S., Nace, R. et al. Meningeal myeloma deposits adversely impact the therapeutic index of an oncolytic VSV. Cancer Gene Ther 20, 616–621 (2013). https://doi.org/10.1038/cgt.2013.63
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cgt.2013.63
Keywords
This article is cited by
-
Chemogenetic ON and OFF switches for RNA virus replication
Nature Communications (2021)
