
Abstract
Serine/threonine kinase family members p21-activated kinases (PAKs) are important regulators of cytoskeletal remodeling and cell motility in mononuclear phagocytic system, but their role in macrophage differentiation and polarization remains obscure. We have shown here that inflammatory stimuli induced PAK1 overexpression in human and murine macrophages. Elevated expression of PAK1 contributed to macrophage M1 polarization and lipopolysaccharide (LPS)-induced endotoxin shock. We further observed that epigenetic loss of microRNA let-7c due to enhancer of zeste homolog 2 (EZH2) upregulation determined PAK1 elevation and inflammatory phenotype in M1 macrophages. EZH2/let-7c/PAK1 axis promotes macrophage M1 polarization via NIK-IKK-NF-κB signaling. Moreover, pharmacological and genetic ablation with EZH2/let-7c/PAK1 axis blunted inflammatory phenotype in M1 macrophages. Critically, either myeloid-restricted PAK1 deletion (PAK1Lyz2cre) or pharmacological and genetic ablation with EZH2/let-7c/PAK1 signal resulted in resistance to LPS-induced endotoxin shock via blunting macrophage M1 polarization. PAK1, therefore, is an essential controller of inflammatory macrophage polarization, regulating immune responses against pathogenic stimuli.
Similar content being viewed by others


Main
Macrophages are key heterogeneous population of immune cells to the initiation, progression, and resolution of inflammation, with diverse functionally polarized subtypes involved at each step.1, 2 Polarized macrophages is often simplified into two broad opposing categories: classical activated macrophages, termed M1, induced by microbial ligands and cytokines such as lipopolysaccharide (LPS) and interferon-γ (IFN-γ) or granulocyte–macrophage colony-stimulating factor (GM-CSF), and alternative activated macrophages, termed M2, as a result of exposure to IL-4, IL-13, IL-10, glucocorticoids, and immune complexes plus TLR ligands.3, 4, 5 While M1 macrophages produce pro-inflammatory mediators such as tumor necrosis factor-α (TNF-α), IL-6, IL-12, IL-23, and NO, and have a role in killing intracellular microorganisms, M2 macrophages produce anti-inflammatory mediators such as IL-4, IL-5, IL-10, and IL-13, and are important in atopic disease, parasite response, and tumor progression.6, 7, 8 The diverse polarized phenotypes of M1–M2 macrophages can, to some extent, be dynamically reversed in vitro and in vivo.1 Moreover, pathology is frequently associated with dynamic changes of macrophage polarization, with M1 macrophages implicated in initiating and sustaining acute inflammation and M2 macrophages associated with resolution or smoldering chronic inflammation.9, 10, 11, 12 Thus, macrophage polarization is an important component of many disease states, including infection, insulin resistance, atherosclerosis, cancer, and autoimmune disease.3, 13, 14, 15 The identification of molecular mechanisms underlying macrophage plasticity and polarization provides a basis for macrophage-centered diagnostic and therapeutic strategies.
A network of signaling molecules, transcription factors, and epigenetic regulators underlies the different forms of macrophage polarization. Canonical IFN regulatory factor (IRF)/STAT signaling pathways are activated by IFNs and TLR signaling to skew macrophage function toward the M1 phenotype (via IRF3, IRF5, IRF8, and STAT1) or by IL-4, IL-13, and IL-10 to skew toward the M2 phenotype (via IRF4, STAT3, and STAT6).13, 16, 17 Suppressors of cytokine signaling 1 (SOCS1), SOCS2, and SOCS3 have been identified as key diametric regulators of macrophage polarization by regulating cytokine–STAT signaling.18, 19 Downstream of, or in parallel with, the IRF/STAT/SOCS pathway, a panel of transcription factors orchestrates diametrical macrophage polarization. The transcription factor NF-κB pathway and MAPK signal are considered crucial orchestrators for inflammatory macrophage polarization.10, 11, 20 The nuclear receptors PPARγ and PPARδ, Krüppel-like factor 4 (KLF4), KLF2, c-Myc, c-Myf, p50 NF-κB homodimers, and HIF-2α are implicated in M2 macrophage polarization.1 Besides, epigenetic regulators and noncoding RNAs also participate in directing macrophage polarization. Histone demethylase jumonji domain containing-3 (JMJD3) and histone deacetylase 3 (HDAC3) have been identified as epigenetic accelerator and brake for M2 macrophage polarization, respectively.17, 21 MicroRNAs (miRNAs) including miR-155, miR-125, miR-146, miR-9, miR-21, miR-147, miR-223, miR-187, miR-378-3p, miR-511-3p, miR-19a-3p, miR-124, and let-7c were recently identified as diverse orchestrators of macrophage polarization.22, 23
The p21-activated kinase (PAK) family of serine/threonine kinases defines an evolutionally conserved crucial convergent signaling nodule that participates in pleiotropic physiological processes including cytoskeleton dynamics and cell motility, survival, mitosis, transcription, and translation.24 On the basis of structural and functional similarities, the six members of PAK family are classified into two groups with three members in each group.25 In contrast to constitutively active group II PAKs (PAK4–6), group I PAKs (PAK1–3) relay signals from Rho GTPase family members cell division cycle 42 and Rac1 to downstream signaling events via activating additional kinases and other effectors by phosphorylating them at specific serine or threonine residues or through protein–protein interactions.26 Group I PAKs have been identified as crucial regulators in immune responses and viral infections.27, 28 The Akt family members of serine/threonine kinases including Akt1 and Akt2 have been found to diametrically orchestrate diverse macrophage polarization via miR-155-mediated C/EBPβ signal regulation.29, 30 Although our previous findings indicated the crucial role of PAK1 in anoikis resistance and hepatoma progression,31 the potential significance of PAKs family in macrophage polarization remains far from fully understood.
In the present study, we found that PAK1 overexpression was induced in macrophages challenged with inflammatory stimuli, and provided evidence for enhancer of zeste homolog 2 (EZH2)-mediated loss of let-7c that determines inflammatory macrophage polarization via PAK1-dependent NF-κB activation. More importantly, we demonstrated that genetic and pharmacological elimination of EZH2/let-7c/PAK1 signal conferred mice with resistance to LPS via blunting macrophage M1 polarization. Our data suggest that activation of PAK1 signals defines commitment to the macrophage lineage by driving inflammatory macrophage polarization and by highlighting the potential for therapeutic interventions via modulation of EZH2/let-7c/PAK1 axis.
Results
Elevated expression of PAK1 in inflammatory polarized macrophages
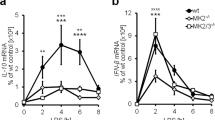
The inflammatory macrophages have long been known to be induced by LPS alone or in concert with cytokines such as IFN-γ or GM-CSF. Treatment of macrophages with well-established inflammatory stimuli (LPS plus IFN-γ) resulted in upregulation of PAK1 mRNA expression compared with no significant changes of PAK2–6 gene expression in RAW264.7 cells (Figure 1a), thioglycollate-elicited mouse peritoneal macrophages (PMs; Figure 1b), and mouse bone marrow-derived macrophages (BMDMs; Figure 1c). Immunoblotting analysis indicated significant elevated PAK1 protein levels in murine BMDMs after LPS plus IFN-γ treatment (Figures 1d and e). Similar effects on PAK1 expression were also obtained in THP1 cells and primary human macrophages treated with LPS plus IFN-γ (Figures 1f and g). To account for possible differences in protocols for the in vitro differentiation of macrophages, we analyzed the amount of PAK1–6 in macrophages treated with GM-CSF and found that these were similar to those in LPS plus IFN-γ-treated cells (Supplementary Figures 1a–e). These results led to the hypothesis that PAK1 elevation induced by inflammatory stimuli might function as a facilitator of inflammatory macrophage polarization.

PAK1 expression is augmented by inflammatory stimuli in macrophages. (a–c) PAK1 mRNA levels were assessed by real-time PCR and normalized to GAPDH in RAW264.7 cells (a), mouse PMs (b), and BMDMs (c) after 12 h of stimulation with LPS (50 ng/ml) plus IFN-γ (20 ng/ml). (d and e) PAK1 protein levels were analyzed by immunoblot and normalized to GAPDH in murine BMDMs after LPS plus IFN-γ stimulation (d). Graphical representation of band intensities was shown in e. (f and g) PAK1 mRNA levels were quantified by real-time PCR and normalized to GAPDH in THP1 cells (f) or GM-CSF-differentiated primary human macrophages (g) stimulated with LPS plus IFN-γ for 12 h. Data are from four independent experiments (a–c and e–g; mean±S.E.M.) or one experiment representative of four independent experiments with similar results (d). ***P<0.001
PAK1 controls M1 macrophage-associated genes
To investigate the role of PAK1 in macrophage activation, we profiled the gene expression of wild-type and PAK1-deficient BMDMs stimulated with LPS plus IFN-γ, which induces the expression of key M1 macrophage-associated proteins, such as TNF-α, IL-6, IL-12, and iNOS. We confirmed efficient deletion of PAK1 in BMDMs from PAK1-deficient mice by assessing the expression of PAK1 mRNA (Supplementary Figure 2a). We found that PAK1 controlled the expression of M1 macrophage-associated genes including TNF-α, IL-6, IL-12a, IL-12b, MCP-1, and Nos2 (Figure 2a). To assess the functional and the physiological relevance of PAK1-mediated regulation of M1 macrophage-associated genes, we examined the in vivo expression of M1-associated cytokines in the myeloid compartment under conditions of inflammation. Real-time PCR and immunoblot analysis confirmed specific ablation of PAK1 expression in macrophages isolated from myeloid-restricted PAK1-deficient (ΦKO) mice, but not from control (ΦWT) mice (Supplementary Figures 2b and c). Endotoxin-induced model of sepsis with a lethal dose of LPS intraperitoneally (i.p.) injection showed that ΦKO mice were resistant to LPS lethality, whereas ΦWT mice succumbed within 30 h (Figure 2b). More importantly, murine serum amounts of inflammatory cytokines including TNF-α, IL-6, IL-12, and MCP-1 were significantly lower in ΦKO mice compared with ΦWT mice after LPS challenge for 6 h (Figure 2c). Similar effects on mRNA levels of aforementioned inflammatory cytokines were observed in murine PMs obtained from ΦKO mice compared with ΦWT mice after LPS challenge for 6 h (Figure 2d). Moreover, inflammatory stimuli-induced inflammatory cytokines production, such as TNF-α, IL-6, IL-12, IL-1β, MCP-1, and nitrite, were alleviated in murine PMs obtained from ΦKO mice compared with ΦWT mice (Figure 2e). These results showed that PAK1 was essential for the expression of genes characteristic of the core M1 macrophage response in vitro and for the manifestation of key myeloid effector functions in vivo.

PAK1 controls the expression of prototypical M1 macrophage-associated genes. (a) TNF-α, IL-6, IL-12a, IL-12b, MCP-1, and Nos2 mRNA levels were quantified by real-time PCR and normalized to GAPDH in BMDMs obtained from PAK1-deficient (PAK1-KO) mice and their wild-type (WT) littermates (controls) and stimulated for 0–6 h (horizontal axes) with LPS (50 ng/ml) plus IFN-γ (20 ng/ml). (b) Kaplan–Meier survival analysis within 50 h for myeloid-specific PAK1-deficienct (ΦKO) mice and wild-type (ΦWT) mice (n=12 per group) after lethal dose LPS challenge with 6 mg/kg i.p. injection. (c) Serum amounts of TNF-α, IL-6, IL-12p40, and MCP-1 were quantified by ELISA in ΦKO mice and ΦWT mice (n=12 per group) after lethal dose LPS challenge for 6 h. (d) Expression levels of TNF-α, IL-6, IL-12b, and MCP-1 were quantified by real-time PCR and normalized to GAPDH in PMs obtained from ΦKO mice and ΦWT mice after LPS challenge for 6 h. (e) Supernatant amounts of TNF-α, IL-6, IL-12p40, IL-1β, MCP-1, and nitrite were quantified by ELISA in PMs obtained from ΦKO and ΦWT mice after LPS plus IFN-γ stimulation for 12 h. Data are representative of six experiments (a; mean±S.E.M.), three independent experiments (e; mean±S.E.M.), or one experiment (b; n=12 mice per group), or are pooled from 8 to 12 independent experiments (c and d). **P<0.01 and ***P<0.001
PAK1 mRNA is a novel bona fide target of let-7c
To further define molecular mechanism for PAK1 elevation induced by inflammatory stimuli, we revealed that 3′UTR of PAK1 gene significantly increased in M1 macrophages (Figure 3a and Supplementary Figure 3a). MiRNAs are a class of noncoding small RNAs that bind to the 3′UTR of target genes, thereby inhibiting their expression through repressing mRNA translation and/or inducing degradation of target gene transcripts.32 We then screened inflammatory stimuli-treated RAW264.7 cells using real-time PCR to identify the possible miRNAs that might modulate PAK1 expression. In a screening for 18 potential miRNAs targeting 3′UTR of PAK1 mRNA, we observed significantly suppressed let-7c RNA levels in inflammatory stimuli-treated RAW264.7 cells (Figure 3b and Supplementary Figure 3b). The PAK1 3′UTR has conserved sites complementary to the let-7c RNA (Figure 3c), leading us to suspect that disrupting let-7c regulation of PAK1 might lead to increased expression of PAK1 in M1 macrophages. Consistent with this idea, introducing let-7c mimics repressed the wild type but not mutant type of PAK1 3′UTR activity in a dose-dependent manner (Figure 3d). To further validate PAK1 as a legitimate target of let-7c, we examined the expression of the endogenous PAK1 when let-7c expression or deletion was manipulated. Introduction of let-7c mimics (Figure 3e and Supplementary Figure 3c) and AS-let-7c oligos (Figure 3f and Supplementary Figure 3d) into RAW264.7 cells significantly reduced and increased PAK1 mRNA levels, respectively. These data strongly suggested that decreased let-7c expression might lead to PAK1 upregulation in M1 macrophages.

PAK1 mRNA is identified as a bona fide target of let-7c in M1 macrophages. (a) Luciferase activities for transfected PAK1 3′UTR were measured and normalized to Renilla luciferase activities in RAW264.7 cells after LPS plus IFN-γ stimulation. (b) Relative levels of potential miRNAs targeting PAK1 mRNA were measured by real-time PCR and normalized to U6 levels in RAW264.7 cells after LPS plus IFN-γ stimulation. (c) Schematic showing putative let-7c binding sites in the wild-type (WT) PAK1 3′UTR (PAK1-3′UTR-WT) sequence and the mutant construct (PAK1-3′UTR-MT) with the putative target sequences mutated. (d) Luciferase activities for transfected PAK1-3′UTR-WT and PAK1-3′UTR-MT were measured and normalized to Renilla luciferase activities at 24 h in HEK293T cells cotransfected with increasing amounts of let-7c mimics. (e and f) PAK1 mRNA levels were assessed by real-time PCR and normalized to GAPDH in RAW264.7 cells transfected with increasing amounts of let-7c mimics (e) and AS-let-7c oligos (f) after stimulated with LPS plus IFN-γ for 12 h. Data are from at least four independent experiments (a, b, and d–f; mean±S.E.M.). *P<0.05, **P<0.01, and ***P<0.001
EZH2 suppresses let-7c expression in inflammatory polarized macrophages
Since previous study unraveled suppressive effect of PcG repressor complex subunit EZH2 on several miRNAs including let-7c,33 we hypothesized that epigenetic repressor EZH2 might have crucial role in M1 stimuli-induced loss of let-7c. Immunoblotting analysis showed substantially increased EZH2 protein levels in RAW264.7, PM, and BMDM cells upon inflammatory stimulation (Figure 4a). To further determine the suppressive role of EZH2 and its catalytic product trimethylation of lysine 27 of histone H3 (H3K27me3) on let-7c, the enriched DNA from the immunoprecipitates was quantified by real-time PCR using primers spanning the let-7c upstream regions (Figure 4b). Enrichment of EZH2 (Figure 4c) and H3K27me3 (Figure 4d) was found to be associated with the let-7c upstream region in RAW264.7, PM, and BMDM cells upon LPS plus IFN-γ stimulation. Moreover, real-time PCR and CHIP experiments showed that specific shRNA against EZH2 (Figure 4e), miR-101 as a well-established miRNA targeting EZH234 (Figure 4f), and 3-deazaneplanocin A (DZNep) as a global histone methylation inhibitor (Figure 4g) restored let-7c expression accompanied with decreased enrichment of H3K27me3 on let-7c upstream region in M1 macrophages. These results identified EZH2 as an epigenetic suppressor for let-7c expression in M1 macrophages.

EZH2 mediates epigenetic suppression of let-7c transcription in M1 macrophages. (a) EZH2 protein levels were analyzed by immunoblot and normalized to GAPDH in RAW264.7 cells, PMs, and BMDMs after stimulated with LPS plus IFN-γ or GM-CSF for 24 h. (b) Schematic showing genomic location of let-7c and the upstream regions used for ChIP-PCR are indicated as short black lines. (c and d) Enrichment of EZH2 (c) and H3K27me3 (d) at indicated locations upstream of let-7c were quantified by ChIP-PCR and normalized to IgG in RAW264.7 cells, PMs, and BMDMs after LPS plus IFN-γ stimulation. (e–g) Expression levels of EZH2 and let-7c (left panel) were quantified by real-time PCR and normalized to GAPDH and U6, respectively, and enrichment of H3K27me3 in upstream region of let-7c (right panel) were quantified by ChIP-PCR and normalized to IgG in LPS plus IFN-γ-treated RAW264.7 cells after EZH2–shRNA introduction (e), miR-101 transfection (f), and DZNep treatment (g). Data are representative results obtained from at least three independent experiments (a and c–g; mean±S.E.M.). *P<0.05, **P<0.01, and ***P<0.001, NS, not significant
EZH2/let-7c/PAK1 axis determines inflammatory macrophage phenotype
We next examined whether EZH2/let-7c/PAK1 signal determines inflammatory phenotype in M1 macrophages. In parallel with substantially elevated let-7c and suppressed PAK1 levels by real-time PCR analysis (Supplementary Figures 4a–c), both EZH2 deletion with EZH2–shRNA and EZH2 blockade with DZNep induced significant decreased inflammatory phenotypic cytokines including TNF-α, IL-6, IL-12, IL-1β, RANTES, and MCP-1 by ELISA analysis in RAW264.7 cells treated with LPS plus IFN-γ (Figure 5a). Moreover, compared with no substantial effect on EZH2 levels, let-7c restoration with let-7c mimics and suppression with AS-let-7c oligos induced, respectively, decreased/increased PAK1 levels by real-time PCR analysis (Supplementary Figures 4d–f) and decreased/increased inflammatory phenotypic cytokines including TNF-α, IL-6, IL-12, IL-1β, RANTES, and MCP-1 by ELISA analysis in RAW264.7 cells treated with LPS plus IFN-γ (Figure 5b). Similar effects on EZH2 and PAK1 levels by real-time PCR analysis (Supplementary Figures 4g–i) and inflammatory phenotypic cytokines by ELISA analysis (Figure 5c) were observed after introduction with a lentiviral Tet-On vector to overexpress inducibly let-7c upon Dox exposure and a lentiviral vector expressing a let-7c ‘sponge’ to suppress let-7c expression, respectively, in RAW264.7 cells treated with LPS plus IFN-γ. Critically, in parallel with no substantial effects on EZH2 and let-7c levels by real-time PCR analysis (Supplementary Figures 4j–l), PAK1 knockdown with PAK1–shRNA, inactivation with kinase dead mutant PAK1-K299R, and inhibition with allosteric inhibitor PAK inhibitor III (IPA3) significantly reversed inflammatory phenotypic cytokines by ELISA analysis in RAW264.7 cells treated with LPS plus IFN-γ (Figure 5d). Collectively, these findings demonstrated that EZH2/let-7c/PAK1 axis orchestrates inflammatory phenotype in M1-polarized macrophages.

EZH2/let-7c/PAK1 axis facilitates macrophage M1 polarization. (a–d) Supernatant amounts of TNF-α, IL-6, IL-12p40, IL-1β, RANTES, and MCP-1 were quantified by ELISA in LPS plus IFN-γ-treated RAW264.7 cells after EZH2–shRNA introduction or DZNep treatment (a), LPS plus IFN-γ-treated RAW264.7 cells after let-7c mimics or AS-let-7c oligos introduction (b), LPS plus IFN-γ-treated RAW264.7 cells after let-7c sponge lentivirus introduction or RAW264.7 Tet-on-let-7c cells with or without Dox treatment (c), and LPS plus IFN-γ-treated RAW264.7 cells after PAK1–shRNA introduction, PAK1-K299R transfection, or IPA3 treatment (d). Data are from four independent experiments (mean±S.E.M.). **P<0.01 and ***P<0.001
PAK1 promotes inflammatory macrophage polarization via NIK-IKK-NF-κB activation
Previous studies indicating PAK1-mediated NF-κB activation35 and well-established role of NF-κB in macrophage polarization36 led us suspect the potential participation of NF-κB signal in PAK1-induced inflammatory macrophage polarization. Luciferase reporter assay showed that PAK1 knockdown with PAK1–shRNA, PAK1 inhibition with IPA3, and PAK1 inactivation with PAK1-K299R reversed NF-κB activation, in contrast to facilitation of NF-κB activation due to PAK1 activation with PAK1-WT and constitutive active mutant PAK1-T423E in RAW264.7 cells challenged with LPS plus IFN-γ (Figures 6a and b). Similar effects of PAK1 inactivation with PAK1-K299R and PAK1 activation with PAK1-WT and PAK1-T423E on TNF-α production were observed by ELISA analysis in RAW264.7 cells challenged with LPS plus IFN-γ (Figure 6c). Moreover, blockade of NF-κB activation with PDTC, Bay 11–7082, and IκBα-SR notably suppressed NF-κB activity and TNF-α production mediated by PAK1 activation with PAK1-T423E (Figures 6d–g). Further analysis of potential roles for upstream molecules of NF-κB signal, such as NIK, MEKK1, IKKα, and IKKβ, in PAK1-mediated NF-κB activation showed that shRNA-mediated knockdown of NIK, IKKα, or IKKβ, but not MEKK1, could pronouncedly block NF-κB activity and TNF-α production mediated by PAK1 activation with PAK1-T423E (Figures 6h and i). The phosphorylation status of nuclear NF-κB determines its association with CBP/p300 or HDAC-1 during NF-κB-dependent transcription.37 ChIP analysis showed that PAK1 inactivation with PAK1-K299R reversed enrichment of epigenetic activators including CBP/p300 and H3K4me3, and epigenetic suppressors comprising HDAC-1, H3K27me3, and H3K9me3, whereas PAK1 activation with PAK1-T423E facilitated enrichment of aforementioned epigenetic regulators on TNF-α upstream regions in RAW264.7 cells incubated with LPS plus IFN-γ (Figures 6j–n). These data suggested that PAK1 might promote inflammatory macrophage polarization via NIK-IKK-NF-κB signal-mediated transcriptional activation of phenotypic markers.

PAK1 fosters macrophage M1 polarization via NIK-IKK-NF-κB activation. (a, b, d, f, and h) Luciferase activities for transfected NF-κB responsive elements were measured and normalized to Renilla luciferase activities in LPS plus IFN-γ-treated RAW264.7 cells after PAK1–shRNA introduction or IPA3 treatment (a), LPS plus IFN-γ-treated RAW264.7 cells after PAK1-WT, PAK1-K299R, or PAK1-T423E introduction (b), PAK1-T423E-transfected RAW264.7 cells after PDTC or Bay 11–7082 treatment (d), PAK1-T423E-transfected RAW264.7 cells after increasing amounts of IκBα superrepressor (IκBα-SR) introduction (f), and PAK1-T423E-transfected RAW264.7 cells after NIK-shRNA, MEKK1-shRNA, IKKα-shRNA, or IKKβ-shRNA introduction (h). (c, e, g, and i) Supernatant amounts of TNF-α were quantified by ELISA in LPS plus IFN-γ-treated RAW264.7 cells after PAK1-WT, PAK1-K299R, or PAK1-T423E introduction (c), PAK1-T423E-transfected RAW264.7 cells after PDTC or Bay 11–7082 treatment (e), PAK1-T423E-transfected RAW264.7 cells after increasing amounts of IκBα superrepressor (IκBα-SR) introduction (g), and PAK1-T423E-transfected RAW264.7 cells after NIK-shRNA, MEKK1-shRNA, IKKα-shRNA, or IKKβ-shRNA introduction (i). (j–n) Enrichment of p300 (j), HDAC-1 (k), H3K27me3 (l), H3K9me3 (m), and H3K4me3 (n) at upstream regions of TNF-α were quantified by ChIP-PCR and normalized to IgG in LPS plus IFN-γ-treated RAW264.7 cells after PAK1-K299R or PAK1-T423E introduction. Data are from four to six independent experiments (a, b, d, f, and h; mean±S.E.M.), from five to six independent experiments (c, e, g, and i; mean±S.E.M.), or are representative results from four independent experiments (j–n; mean±S.E.M.). *P<0.05, **P<0.01, and ***P<0.001; NS, not significant
Blockade of EZH2/let-7c/PAK1 signal confers LPS resistance
To define the crucial role of myeloid EZH2/let-7c/PAK1 signal for inflammatory macrophage polarization in vivo, LPS lethality and serum amounts of inflammatory cytokines were monitored after EZH2/let-7c/PAK1 signal intervention in endotoxin-induced murine sepsis model. Regulatory effect on EZH2, let-7c, and PAK1 expression in PMs obtained from DZNep- or IPA3-treated mice were confirmed by real-time PCR analysis (Figures 7a and e). Moreover, DZNep- or IPA3-treated mice were resistant to lethal dose of LPS treatment, whereas control mice succumbed within 24 h (Figures 7b and f). Murine serum amounts of inflammatory cytokines including TNF-α, IL-6, IL-12, IL-1β, RANTES, and MCP-1 were significantly lower in DZNep- or IPA3-treated mice compared with control mice after LPS challenge for 6 h (Figures 7c and g). Besides, inflammatory stimuli (LPS plus IFN-γ)-induced cytokines production, such as TNF-α, IL-6, IL-12, IL-1β, MCP-1, and nitrite, were alleviated in murine PMs obtained from DZNep- or IPA3-treated mice compared with control mice (Figures 7d and h). These data indicated that EZH2/let-7c/PAK1 signal intervention could lead to LPS resistance via blunting macrophage M1 polarization. Together, we propose a model for the regulation of the polarization of M1 macrophages through PAK1-dependent NF-κB signaling pathway (Supplementary Figure 5).

Blockade of EZH2/let-7c/PAK1 signal confers LPS resistance via blunting macrophage M1 polarization. (a and e) Expression levels of EZH2, let-7c, and PAK1 were quantified by real-time PCR and normalized to GAPDH and U6, respectively, in PMs obtained from DZNep- (a) and IPA3-treated (e) mice (n=12 mice per group). (b and f) Kaplan–Meier survival analysis within 50 h for DZNep- (b) and IPA3-treated (f) mice (n=12 mice per group) with lethal dose LPS challenge. (c and g) Serum amounts of TNF-α, IL-6, IL-12p40, IL-1β, RANTES, and MCP-1 were quantified by ELISA in DZNep- (c) and IPA3-treated (g) mice (n=12 mice per group) after lethal dose LPS challenge for 6 h. (d and h) Supernatant amounts of TNF-α, IL-6, IL-12p40, IL-1β, MCP-1, and nitrite were quantified by ELISA in PMs obtained from DZNep- (d) and IPA3-treated (h) mice (n=12 mice per group) after LPS plus IFN-γ stimulation for 12 h. Data are representative of four experiments (a and e; mean±S.E.M.), one experiment (b and f; n=12 mice per group), pooled from 6 to 12 independent experiments (c and g), or three independent experiments (d and h; mean±S.E.M.). **P<0.01 and ***P<0.001; NS, not significant
Discussion
Our findings herein indicate that PAK1 is induced in macrophages activated by inflammatory stimuli and controls macrophage inflammatory polarization, and further provide evidence for EZH2-mediated epigenetic loss of let-7c that determines inflammatory macrophage polarization via PAK1-dependent NIK-IKK-NF-κB signal activation. More importantly, our data also show that EZH2/let-7c/PAK pharmacological intervention with DZNep or IPA3 treatment conferred mice with LPS resistance via blunting macrophage M1 polarization. In addition to unravel molecular signals underlying inflammatory macrophage polarization, our results have potential clinical significance for therapy against endotoxin-induced sepsis and inflammation. Besides, we have also observed additional pivotal significance of myeloid PAK1 in suppressing M2 macrophage polarization (unpublished data), which was excluded from this study focusing on delineation of molecular mechanism for M1 macrophage polarization.
As a crucial family of serine/threonine kinases, PAKs have dozens of known substrates whose phosphorylation affects numerous cellular processes, including cytoskeletal organization, cell cycle progression, and cell survival, as well as significant non-kinase-related scaffold effects. Among group I PAK family members, PAK1 and PAK2, but not PAK3, are expressed at high levels in leukocytes and have important functions in a variety of white blood cells, including T cells, neutrophils, macrophages, and mast cells.27 T-cell receptor-mediated activation of Erk and NFAT depends on PAK1 activity that acts downstream of an Nck/Vav/SLP-76 signaling complex.38 PAK1 activity is required for chemotaxis of leukocytes in response to the chemokine CXCL1.39 Using PAK1−/− bone marrow-derived mast cells, Allen et al.40 demonstrated that PAK1 is required for IgE-mediated calcium influx and degranulation. Macrophages derived from PAK1−/− mice also display reduced Erk activation in response to growth factors as well as defects in the stability of lamellipodia.41 Consistent with previous study indicating that endotoxin LPS is a potent activator of PAK1 kinase activity,35 our present study also implicates that PAK1 expression is induced by inflammatory stimuli (LPS plus IFN-γ and GM-CSF) in murine and human macrophages, which leads us to suspect the possible role of PAK1 in inflammatory macrophage polarization.
Furthermore, our data identified PAK1 as a bona fide target of let-7c in inflammatory polarized macrophages, indicating let-7c as a novel epigenetic regulator for PAK1. Besides, our results clarify the potential significance of PAK1 kinase activity in sustaining inflammatory polarized phenotype through genetic deletion of PAK1 expression and pharmacological inhibition PAK1 activity. We also identified NIK-IKK-NF-κB signal as essential module relaying PAK1 activity to inflammatory polarized phenotype. More importantly, we found that myeloid-restricted PAK1 deficiency confers mice with resistance to endotoxin-induced sepsis, suggesting pivotal role of PAK1 in inflammatory response in mice challenged with M1-like pathogenic stimuli. Critically, we also observed that IPA3-treated mice exhibit resistance to endotoxin-induced sepsis and alleviated inflammatory macrophage polarization, identifying IPA3 as a potential therapeutic drug against endotoxin-induced sepsis and inflammation.
Previous studies have suggested the crucial regulatory role of miRNAs including miR-155, miR-125, miR-146, miR-9, miR-21, miR-147, miR-223, miR-187, miR-378-3p, miR-511-3p, miR-19a-3p, miR-124, and let-7c in macrophage polarization.22, 23 Our present findings reveal the crucial significance of let-7c in inflammatory macrophage polarization, which is concordant with previous study indicating that let-7c could suppress polarization of macrophages to the M1 phenotype and enhances M2 polarization via targeting C/EBP-delta.42 Moreover, our study indicates PAK1 as another authentic target of let-7c and upstream kinase for NIK-IKK-NF-κB activation and following inflammatory macrophage polarization. Collectively, let-7c might be used as a crucial rheostat for diametrical macrophage M1–M2 polarized phenotype conversion. Besides, our current results implicate that let-7c mimics could be used as a potential therapeutic agent against myeloid PAK1 overactivation in endotoxin-induced sepsis and inflammation. Although the targeting of miRNAs for therapeutic purposes is at an early stage, given the crucial role of let-7c in the control of inflammation, and, in particular, in macrophage activation, they remain of interest for future drug development.
Epigenetic control of gene expression is crucial for cellular differentiation and development.43, 44 Emerging evidence suggests an important role for epigenetic mechanisms including DNA and histone modifications in modulating and transmitting signals during macrophage polarization and reprogramming.45, 46 Epigenetic landscape of macrophages including DNA methylation and chromatin modifications on potential target genes determines accessibility for binding with signaling transcription factors such as NF-κB and STAT3 induced by extracellular stimuli and thus orchestrating diverse functional phenotypes. Previous study observed that IL-4 induces upregulation of the JMJD3 in mouse macrophages that alters chromatin modifications to promote expression of M2 genes and inhibit M1 genes.17 Our present study reveals that EZH2 is induced by M1-like stimuli and mediates epigenetic loss of let-7c in inflammatory polarized macrophages. However, molecular signals accounting for EZH2 induction in response to M1-like stimuli need further investigations. Our current data also reveal that both pharmacologic and genetic inhibition of EZH2 could lead to let-7c-mediated PAK1 repression and macrophage inflammatory phenotype reversion. Thus, we have shown here that PAK1-dependent NF-κB signaling pathway was required for macrophage inflammatory polarization and have designated the EZH2/let-7c/PAK1 regulatory axis as a new target for therapeutic intervention to suppress inflammation that may be applicable to the treatment of inflammatory disorders.
Materials and Methods
Mice
B6.129P2-Lyz2tm1(cre)Ifo/J mice were purchased from Jackson Laboratories (Bar Harbor, ME, USA). PAK1fl/fl mice were constructed by homologous recombination in embryonic stem cells using standard methods as described previously.47 Mice with myeloid cell-specific deletion of PAK1 (PAK1Lyz2cre) were littermates obtained from crosses of B6.129P2-Lyz2tm1(cre)Ifo/J mice with PAK1fl/fl mice. Mice with inducible deletion of PAK1 (PAK1fl/flMx1-Cre) were generated by crossing of PAK1fl/fl mice to mice expressing a Mx1 promoter-driven transgene encoding Cre on the C57/BL6J background. Littermates with an PAK1fl/flMx1-Cre or PAK1+/+ Mx1-Cre genotype were given i.p. injection of poly(I:C) at a dose of 200 μg per mouse three times in 5 days to induce deletion, and 2 weeks later mice were used for experiments. For all in vitro experiments involving PAK1-deficient macrophages, cells were derived from PAK1fl/flMx1-Cre mice where substantial deletion of PAK1 (∼80%) was observed; the conditional deletion was controlled for expression of Cre and genetic background through the use of cells from PAK1+/+ Mx1-Cre mice. C57BL/6J mice were purchased from Experimental Animal Center of Chinese Academy of Sciences (Shanghai, China). Mice were specific pathogen free and used at 6–8 weeks. Experiments were sanctioned and approved by the Ethic Review Committee for Shanghai Medical College of Fudan University.
Cell line and macrophage isolation
RAW264.7, THP1, and human embryonic kidney cell line 293T cells were originally obtained from ATCC. Murine PMs and BMDMs were isolated as described previously.48 Human monocytes were purified as described previously.49 Polarization studies were performed using macrophages incubated with 50 ng/ml LPS (Invivogen, San Diego, CA, USA) plus 20 ng/ml IFN-γ or 50 ng/ml GM-CSF (BD Biosciences, San Jose, CA, USA) for 24 h.
Gene expression analysis
Total RNA was extracted with TRIzol reagent (Invitrogen Life Technologies, Gaithersburg, MD, USA) according to instructions. The cDNA was synthesized with PrimeScript RT reagent kit (Takara Bio, Shiga, Japan). The expression of the genes was quantified by real-time PCR using a Lightcycler480 and SYBR Green system (Roche Diagnostic Systems, Somerville, NJ, USA) following the manufacturer’s protocol.50
Immunoblotting and immunoprecipitation
Protein extraction from cultured cells or tissues, immunoblotting, and immunoprecipitation were carried out as previously described.31, 51 Antibodies used here included those against GAPDH, PAK1, EZH2, H3K27me3, p300, HDAC-1, H3K9me3, and H3K4me3 (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Chromatin immunoprecipitation was carried out with the ChIP Assay Kit (Millipore, Billerica, MA, USA) according to instructions. The percentage of the bound DNA was quantified against the original DNA input by PCR analysis.
Luciferase reporter assay
The luciferase reporter plasmids containing PAK1 3′UTR-WT and PAK1 3′UTR-MT were constructed in pGL3 vector (Promega, Madison, WI, USA). NF-κB luciferase reporter plasmid was obtained from Stratagene (La Jolla, CA, USA). Luciferase reporter assay was performed as described previously.52
Cytokine analysis
TNF-α, IL-6, IL-12, IL-1β, RANTES, and MCP-1 supernatant and serum concentrations were determined by ELISA according to manufacturer’s instructions (R&D Systems, Minneapolis, MN, USA). Nitrite derived from NO was determined with the Griess Reagent System (Promega) in macrophage-conditioned medium according to the manufacturer’s instructions.
let-7c Tet-on and sponge system
let-7c was subcloned into the p-tetracycline response element (pTRE)-tight vector (Clontech, Palo Alto, CA, USA). RAW264.7 were infected with pRevTet-On (Clontech) virus and selected on 500 mg/ml G418 to generate Tet-On cells. RAW264.7 Tet-On cells were cotransfected with pTRE-tight let-7c and a hygromycin resistance gene (Clontech). Positive clones were selected in the presence of G418 and hygromycin, and the inducibility of let-7c was then determined by real-time PCR analysis as described previously.53, 54 Genomic sequence spanning the mouse let-7c coding region was cloned into the lentiviral vector pCDF1-CMV-MCS2-EF1-copGFP downstream of the cytomegalovirus promoter. A miRNA sponge experimental approach was used for suppression of let-7c according to the manufacturer’s instructions.55, 56 Sequence encoding its specific inhibitor (sponge) was cloned into the same vector. Virus was produced and target cells were infected according to the user’s manual (System Biosciences, Mountain View, CA, USA). The efficiency of transduction was ∼40–60%.
Endotoxin-induced model of sepsis and in vivo inhibitor treatment
Sepsis was induced in mice by injecting 6 mg/kg Escherichia coli-derived ultrapure LPS (Invivogen) or PBS (not shown) i.p. Mice were culled immediately at a humane end point noted by loss of self-righting and insensitivity to touch. Survival after LPS was monitored. In some experiments, mice were treated with dimethylsulfoxide plus 6 mg/kg LPS, 1 mg/kg DZNep (Cayman Chemical, Ann Arbor, MI, USA) plus 6 mg/kg LPS, or 4 mg/kg IPA3 (Sigma-Aldrich, St. Louis, MO, USA) plus 6 mg/kg LPS i.p.
Statistical analysis
Comparisons used one-way ANOVA and Tukey post hoc test or utilized two-way ANOVA and Bonferonni post hoc test. Kaplan–Meier graph and log-rank test determined significance of survival. Data were analyzed by GraphPad Prism v5 (GraphPad Software, San Diego, CA, USA). The statistical significance level was set as *P<0.05, **P<0.01, and ***P<0.001.
Abbreviations
- PAKs:
-
p21-activated kinases
- miRNA:
-
microRNA
- EZH2:
-
enhancer of zeste homolog 2
- IRF:
-
IFN regulatory factor
- SOCS:
-
suppressors of cytokine signaling
- HDAC:
-
histone deacetylase
- PM:
-
peritoneal macrophages
- BMDM:
-
bone marrow-derived macrophage
- H3K27me3:
-
trimethylation of lysine 27 of histone H3
- DZNep:
-
3-Deazaneplanocin A
- IPA3:
-
PAK-activated kinase inhibitor III
References
Sica A, Mantovani A . Macrophage plasticity and polarization: in vivo veritas. J Clin Invest 2012; 122: 787–795.
Han MS, Jung DY, Morel C, Lakhani SA, Kim JK, Flavell RA et al. JNK expression by macrophages promotes obesity-induced insulin resistance and inflammation. Science 2013; 339: 218–222.
Mosser DM, Edwards JP . Exploring the full spectrum of macrophage activation. Nat Rev Immunol 2008; 8: 958–969.
Gordon S, Martinez FO . Alternative activation of macrophages: mechanism and functions. Immunity 2010; 32: 593–604.
Tomasini R, Secq V, Pouyet L, Thakur AK, Wilhelm M, Nigri J et al. TAp73 is required for macrophage-mediated innate immunity and the resolution of inflammatory responses. Cell Death Differ 2013; 20: 293–301.
Van Dyken SJ, Locksley RM . Interleukin-4- and interleukin-13-mediated alternatively activated macrophages: roles in homeostasis and disease. Annu Rev Immunol 2013; 31: 317–343.
Wynn TA, Chawla A, Pollard JW . Macrophage biology in development, homeostasis and disease. Nature 2013; 496: 445–455.
Cavnar MJ, Zeng S, Kim TS, Sorenson EC, Ocuin LM, Balachandran VP et al. KIT oncogene inhibition drives intratumoral macrophage M2 polarization. J Exp Med 2013; 210: 2873–2886.
Martinez FO, Helming L, Gordon S . Alternative activation of macrophages: an immunologic functional perspective. Annu Rev Immunol 2009; 27: 451–483.
Zhang W, Xu W, Xiong S . Blockade of Notch1 signaling alleviates murine lupus via blunting macrophage activation and M2b polarization. J Immunol 2010; 184: 6465–6478.
Zhang W, Xu W, Xiong S . Macrophage differentiation and polarization via phosphatidylinositol 3-kinase/Akt-ERK signaling pathway conferred by serum amyloid P component. J Immunol 2011; 187: 1764–1777.
O'Sullivan T, Saddawi-Konefka R, Vermi W, Koebel CM, Arthur C, White JM et al. Cancer immunoediting by the innate immune system in the absence of adaptive immunity. J Exp Med 2012; 209: 1869–1882.
Xu H, Zhu J, Smith S, Foldi J, Zhao B, Chung AY et al. Notch-RBP-J signaling regulates the transcription factor IRF8 to promote inflammatory macrophage polarization. Nat Immunol 2012; 13: 642–650.
Zhang W, Zhou Q, Xu W, Cai Y, Yin Z, Gao X et al. DNA-dependent activator of interferon-regulatory factors (DAI) promotes lupus nephritis by activating the calcium pathway. J Biol Chem 2013; 288: 13534–13550.
Fong CH, Bebien M, Didierlaurent A, Nebauer R, Hussell T, Broide D et al. An antiinflammatory role for IKKbeta through the inhibition of "classical" macrophage activation. J Exp Med 2008; 205: 1269–1276.
Krausgruber T, Blazek K, Smallie T, Alzabin S, Lockstone H, Sahgal N et al. IRF5 promotes inflammatory macrophage polarization and TH1-TH17 responses. Nat Immunol 2011; 12: 231–238.
Satoh T, Takeuchi O, Vandenbon A, Yasuda K, Tanaka Y, Kumagai Y et al. The Jmjd3-Irf4 axis regulates M2 macrophage polarization and host responses against helminth infection. Nat Immunol 2010; 11: 936–944.
Serezani CH, Lewis C, Jancar S, Peters-Golden M . Leukotriene B4 amplifies NF-kappaB activation in mouse macrophages by reducing SOCS1 inhibition of MyD88 expression. J Clin Invest 2011; 121: 671–682.
Spence S, Fitzsimons A, Boyd CR, Kessler J, Fitzgerald D, Elliott J et al. Suppressors of cytokine signaling 2 and 3 diametrically control macrophage polarization. Immunity 2013; 38: 66–78.
Smale ST . Selective transcription in response to an inflammatory stimulus. Cell 2010; 140: 833–844.
Mullican SE, Gaddis CA, Alenghat T, Nair MG, Giacomin PR, Everett LJ et al. Histone deacetylase 3 is an epigenomic brake in macrophage alternative activation. Genes Dev 2011; 25: 2480–2488.
Alam MM, O'Neill LA . MicroRNAs and the resolution phase of inflammation in macrophages. Eur J Immunol 2011; 41: 2482–2485.
Squadrito ML, Etzrodt M, De Palma M, Pittet MJ . MicroRNA-mediated control of macrophages and its implications for cancer. Trends Immunol 2013; 34: 350–359.
Kumar R, Gururaj AE, Barnes CJ . p21-activated kinases in cancer. Nat Rev Cancer 2006; 6: 459–471.
Bokoch GM . Biology of the p21-activated kinases. Annu Rev Biochem 2003; 72: 743–781.
Molli PR, Li DQ, Murray BW, Rayala SK, Kumar R . PAK signaling in oncogenesis. Oncogene 2009; 28: 2545–2555.
Pacheco A, Chernoff J, Group I . p21-activated kinases: emerging roles in immune function and viral pathogenesis. Int J Biochem Cell Biol 2010; 42: 13–16.
Van den Broeke C, Radu M, Chernoff J, Favoreel HW . An emerging role for p21-activated kinases (Paks) in viral infections. Trends Cell Biol 2010; 20: 160–169.
Wei Y, Nazari-Jahantigh M, Chan L, Zhu M, Heyll K, Corbalan-Campos J et al. The microRNA-342-5p fosters inflammatory macrophage activation through an Akt1- and microRNA-155-dependent pathway during atherosclerosis. Circulation 2013; 127: 1609–1619.
Arranz A, Doxaki C, Vergadi E, Martinez de la Torre Y, Vaporidi K, Lagoudaki ED et al. Akt1 and Akt2 protein kinases differentially contribute to macrophage polarization. Proc Natl Acad Sci USA 2012; 109: 9517–9522.
Xu J, Liu H, Chen L, Wang S, Zhou L, Yun X et al. Hepatitis B virus X protein confers resistance of hepatoma cells to anoikis by up-regulating and activating p21-activated kinase 1. Gastroenterology 2012; 143: 199–212.
Stefani G, Slack FJ . Small non-coding RNAs in animal development. Nat Rev Mol Cell Biol 2008; 9: 219–230.
Au SL, Wong CC, Lee JM, Fan DN, Tsang FH, Ng IO et al. Enhancer of zeste homolog 2 epigenetically silences multiple tumor suppressor microRNAs to promote liver cancer metastasis. Hepatology 2012; 56: 622–631.
Varambally S, Cao Q, Mani RS, Shankar S, Wang X, Ateeq B et al. Genomic loss of microRNA-101 leads to overexpression of histone methyltransferase EZH2 in cancer. Science 2008; 322: 1695–1699.
Frost JA, Swantek JL, Stippec S, Yin MJ, Gaynor R, Cobb MH . Stimulation of NFkappa B activity by multiple signaling pathways requires PAK1. J Biol Chem 2000; 275: 19693–19699.
Ivashkiv LB . Inflammatory signaling in macrophages: transitions from acute to tolerant and alternative activation states. Eur J Immunol 2011; 41: 2477–2481.
Zhong H, May MJ, Jimi E, Ghosh S . The phosphorylation status of nuclear NF-kappa B determines its association with CBP/p300 or HDAC-1. Mol Cell 2002; 9: 625–636.
Wu JN, Koretzky GA . The SLP-76 family of adapter proteins. Semin Immunol 2004; 16: 379–393.
Wang D, Sai J, Carter G, Sachpatzidis A, Lolis E, Richmond A . PAK1 kinase is required for CXCL1-induced chemotaxis. Biochemistry 2002; 41: 7100–7107.
Allen JD, Jaffer ZM, Park SJ, Burgin S, Hofmann C, Sells MA et al. p21-activated kinase regulates mast cell degranulation via effects on calcium mobilization and cytoskeletal dynamics. Blood 2009; 113: 2695–2705.
Smith SD, Jaffer ZM, Chernoff J, Ridley AJ . PAK1-mediated activation of ERK1/2 regulates lamellipodial dynamics. J Cell Sci 2008; 121: 3729–3736.
Banerjee S, Xie N, Cui H, Tan Z, Yang S, Icyuz M et al. MicroRNA let-7c regulates macrophage polarization. J Immunol 2013; 190: 6542–6549.
Smale ST, Tarakhovsky A, Natoli G . Chromatin contributions to the regulation of innate immunity. Annu Rev Immunol 2014; 32: 489–511.
Tarakhovsky A . Tools and landscapes of epigenetics. Nat Immunol 2010; 11: 565–568.
Takeuch O, Akira S . Epigenetic control of macrophage polarization. Eur J Immunol 2011; 41: 2490–2493.
Ivashkiv LB . Epigenetic regulation of macrophage polarization and function. Trends Immunol 2013; 34: 216–223.
Liu W, Zi M, Naumann R, Ulm S, Jin J, Taglieri DM et al. Pak1 as a novel therapeutic target for antihypertrophic treatment in the heart. Circulation 2011; 124: 2702–2715.
Zhang X, Goncalves R, Mosser DM . The isolation and characterization of murine macrophages. Curr Protoc Immunol 2008; 83: 14.1.1–14.1.14.
Agostini L, Martinon F, Burns K, McDermott MF, Hawkins PN, Tschopp J . NALP3 forms an IL-1beta-processing inflammasome with increased activity in Muckle-Wells autoinflammatory disorder. Immunity 2004; 20: 319–325.
Liu H, Xu J, Zhou L, Yun X, Chen L, Wang S et al. Hepatitis B virus large surface antigen promotes liver carcinogenesis by activating the Src/PI3K/Akt pathway. Cancer Res 2011; 71: 7547–7557.
Wang S, Zhu Y, He H, Liu J, Xu L, Zhang H et al. Sorafenib suppresses growth and survival of hepatoma cells by accelerating degradation of enhancer of zeste homolog 2. Cancer Sci 2013; 104: 750–759.
Xu J, Yun X, Jiang J, Wei Y, Wu Y, Zhang W et al. Hepatitis B virus X protein blunts senescence-like growth arrest of human hepatocellular carcinoma by reducing Notch1 cleavage. Hepatology 2010; 52: 142–154.
Liu J, Liu H, Zhang W, Wu Q, Liu W, Liu Y et al. N-acetylglucosaminyltransferase V confers hepatoma cells with resistance to anoikis through EGFR/PAK1 activation. Glycobiology 2013; 23: 1097–1109.
Kumar MS, Erkeland SJ, Pester RE, Chen CY, Ebert MS, Sharp PA et al. Suppression of non-small cell lung tumor development by the let-7 microRNA family. Proc Natl Acad Sci USA 2008; 105: 3903–3908.
Ebert MS, Neilson JR, Sharp PA . MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells. Nat Methods 2007; 4: 721–726.
Lee YT, de Vasconcellos JF, Yuan J, Byrnes C, Noh SJ, Meier ER et al. LIN28B-mediated expression of fetal hemoglobin and production of fetal-like erythrocytes from adult human erythroblasts ex vivo. Blood 2013; 122: 1034–1041.
Acknowledgements
This work was supported by grants from National Key Projects for Infectious Disease of China (2012ZX10002-012), National Natural Science Foundation of China (31100629, 31270863, 81472227, 81471621), Program for New Century Excellent Talents in University (NCET-13-0146), and Shanghai Rising-Star Program (13QA1400300).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by R De Maria
Supplementary Information accompanies this paper on Cell Death and Differentiation website
Supplementary information
Rights and permissions
About this article

Cite this article
Zhang, W., Liu, H., Liu, W. et al. Polycomb-mediated loss of microRNA let-7c determines inflammatory macrophage polarization via PAK1-dependent NF-κB pathway. Cell Death Differ 22, 287–297 (2015). https://doi.org/10.1038/cdd.2014.142
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cdd.2014.142
This article is cited by
-
SUZ12 inhibition attenuates cell proliferation of glioblastoma via post-translational regulation of CDKN1B
Genes & Genomics (2023)
-
MicroRNAs in kidney injury and disease
Nature Reviews Nephrology (2022)
-
Characterization of the microRNA transcriptomes and proteomics of cochlear tissue-derived small extracellular vesicles from mice of different ages after birth
Cellular and Molecular Life Sciences (2022)
-
Inhibition of EZH2 prevents acute respiratory distress syndrome (ARDS)-associated pulmonary fibrosis by regulating the macrophage polarization phenotype
Respiratory Research (2021)
-
MicroRNA: Could It Play a Role in Bovine Endometritis?
Inflammation (2021)
