
Abstract
Aim:
To investigate the mechanisms by which berberine suppressed the proliferation of human multiple myeloma cells.
Methods:
Human U266 multiple myeloma cell line was tested. Cell proliferation, apoptosis, ultramicrostructure and secretion function were examined using Cell Counting Kit-8 (CCK8), flow cytometry (FCM), electron and fluorescence microscopy, as well as ELISA assay. The microRNAs (miRs) and transcription factors in U266 cells were detected using arrays and verified by qRT-PCR. EMSA and luciferase assays were used to verify the p65-dependent transactivation of miR-21 gene.
Results:
Treatment of U266 cells with berberine (40−160 μmol/L) suppressed cell proliferation and IL-6 secretion in dose- and time-dependent manners. Meanwhile, berberine dose-dependently induced ROS generation, G2/M phase arrest and apoptosis in U266 cells, and decreased the levels of miR-21 and Bcl-2. Overexpression of miR-21 counteracted berberine-induced suppression of cell proliferation and IL-6 secretion. In U266 cells treated with berberine (80 μmol/L), the activity of NF-κB was decreased by approximately 50%, followed by significant reduction of miR-21 level. berberine (80−160 μmol/L) increased the level of Set9 (lysine methyltransferase) by more than 2-fold, caused methylation of the RelA subunit, which inhibited NF-κB nuclear translocation and miR-21 transcription. In U266 cells treated with berberine (80 μmol/L), knockdown of Set9 with siRNAs significantly increased NF-κB protein level accompanying with a partial recovery of proliferation.
Conclusion:
In U266 cells, berberine suppresses NF-κB nuclear translocation via Set9-mediated lysine methylation, leads to decrease in the levels miR21 and Bcl-2, which induces ROS generation and apoptosis.
Similar content being viewed by others

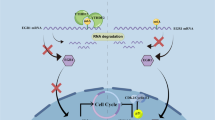

Introduction
Multiple myeloma is one of the most common hematological carcinomas in the aged, and most diagnosed patients often have multiple organ disorders and dyscrasia from both carcinoma and chemotherapy1. In recent decades, various compounds from traditional Chinese medicine have been studied for their anticancer potential. For those compounds with anticancer potential, it is necessary to understand the mechanisms of their anticancer effects, which will guide the development of new cancer therapies. Berberine is a naturally occurring isoquinoline alkaloid that can be extracted from many medicinal herbs, such as Hydrastis canadensis (goldenseal), Cortex phellodendri (Huangbai) and Rhizoma coptidis (Huanglian). Berberine exhibits various pharmacological activities, such as anti-oxidant2, anti-inflammatory3, and anticancer4,5 activities. Moreover, berberine can sensitize human cancer cells to ionizing radiation6 or chemotherapy7. The effect of berberine on leukemia and lymphoma has also been recently studied using both in vivo and in vitro approaches8,9,10. A recent study indicated that berberine downregulates both the mRNA and protein levels of IL-6, which is a key factor in the proliferation of multiple myeloma11. In addition, berberine has been shown to have anti-cachectic and anti-dyscrasia effects in nude mice bearing human esophageal cancer cells12. Therefore, we undertook a detailed study of the sensitivity of multiple myeloma cells to berberine. MicroRNAs (miRs) can lead to either positive or negative regulation at a variety of levels depending on the specific miR, the target base pair interactions and the cofactors that recognize miRs. To further elucidate the mechanism by which berberine affects multiple myeloma, we examined the the miR profile of multiple myeloma cells before and after berberine treatment.
Materials and methods
Materials
Berberine, dimethylsulfoxide and MTT were obtained from Sigma (St Louis, MO, USA). RPMI-1640 medium, fetal bovine serum (FBS) and other cell culture reagents were obtained from Gibco BRL Life Technologies (Grand Island, NY, USA). The following reagents were purchased from Cell Signaling Technology (Danvers, MA, USA): propidium iodide; anti-Bcl-2, anti-Bax, anti-caspase-3, anti-Caspase-9, and anti-PUMA monoclonal antibodies; and secondary antibodies. The following reagents were purchased from Molecular Probes (Eugene, OR, USA): 2′,7′-dichlorofluorescein (DCF), Annexin V and the Fluo-4 NW Calcium Assay kit. The RT-PCR Kit and Trizol were purchased from Invitrogen (Eugene, OR, USA). The mirVANATM miRNA Isolation and Labeling Kit was purchased from Ambion (Austin, TX, USA), and the microRNA detection chip was purchased from Weixin (Shenzhen, China). The miR21 detection kit was purchased from Jima (Shanghai, China).
Cell lines and cell culture
The U266 multiple myeloma cell line was kindly provided by Prof Jie Jin (Department of Hematology, The First Affiliate Hospital of Zhejiang University, Hangzhou, China). The cells were cultured in RPMI medium containing 25 mmol/L HEPES, 10% FBS, 0.05 mmol/L 2-mercaptoethanol, 1 mmol/L sodium pyruvate, 2 mmol/L L-glutamine, 100 U penicillin/mL, and 50 U streptomycin/mL at 37 °C and in 5% CO2. All experiments were performed in triplicate and repeated at least three times.
Cell viability measurement by CCK8 after berberine treatment
U266 cells were plated in 96-well plates at a density of 2×104 cells/well and grown for 24, 48, and 72 h at 37 °C in 5% CO2. The cells were treated with 20 μL of CCK8, and absorbance was recorded at 450 nm. The effect of berberine (0, 40, 80, 120, and 160 μmol/L) on cell viability was assessed as percent cell viability as compared to vehicle-treated control cells, which were arbitrarily assigned 100% viability.
Measurement of IL-6 secretion by ELISA
Levels of IL-6 in cell viability culture supernatants were measured using an enzyme immunoassay (R&D systems, Minneapolis, USA). Briefly, capture antibodies against IL-6 were coated overnight on 96-well ELISA plates, and the plates were blocked using 0.15 mol/L PBS (pH 7.2) with 1% BSA. Diluted culture supernatant was then added to the pre-blocked plates and incubated for 2 h. Bound IL-6 was then detected using a biotinylated anti-IL-6 antibody and streptavidin-HRP (horseradish peroxidase) conjugate. Tetramethylbenzidine was used as the substrate, and absorbance was measured at 450 nm using an ELISA reader (Tecan, Germany). The minimum detection limit of the assays was 9.3 pg/mL.
Ultramicrostructure change of U266 cells after berberine treatment using transmission electron microscopy (TEM) and scanning electron microscopy (SEM)
U266 cells treated with 80 μmol/L berberine for 24 h were centrifuged at 2000×g for 5 min and then prefixed in 2% glutaraldehyde at room temperature for 2 d. The cell were washed three times with 0.1 mol/L PBS (pH 7.2), postfixed in 1% aqueous OsO4 for 2 h, and then washed again three times with 0.1 mol/L PBS (pH 7.2). The resulting samples were placed in isoamyl acetate for 20 min before being dehydrated in a series of ethanol/water washes (25%, 50%, 75%, two 95%, and three 100% ethanol washes). Finally, samples were dried using a critical point drying apparatus, firmly mounted, and sputter coated with a thin layer of gold before being examined under a HITACHI S-570 electron microscope.
Measurement of apoptosis, cell cycle and reactive oxygen species generation
U266 cells were plated in 6-well plates at a density of 2×106 cells/well and grown for 24 h. Various amounts of berberine were added to the cells to final concentrations of 0, 40, 80, 120, and 160 μmol/L. The cells from each treatment were collected and divided into four samples. With the first sample, the enumeration of apoptotic cells was performed using Annexin V-FITC and PI (BioVision, USA). The cells were gently vortexed and resuspended in binding buffer at a concentration of 3×106/mL, and 100 μL of the cell suspension was added to 5 μL of Annexin V-FITC and 10 μL of PI. The samples were mixed for 15 min in the dark at room temperature, and 400 μL of PBS was then added to the solution. A FACScan (Becton Dickinson, San Jose, CA, USA) was used to count cells (1×103) at an excitation wavelength of 490 nm. CellQuest software was used for data collection and processing. With the second sample, cells were resuspended in a buffer containing 2 μg/mL Hoechst 33342 and PI at 28 °C for 30 min and washed three times (5 min each) with PBS (pH 7.2) before detecting apoptosis by fluorescence microscopy. With the third sample, cells were fixed in 70% ethanol at 4 °C for at least 24 h, centrifuged at 2000 r/min for 5 min and washed with PBS for twice, then RNase A (100 μL) was then added to the cells, and placed in a 37 °C water bath for 30 min. PI (400 μL) was then added to the cells, and the cells were protected from light for 30 min at 4 °C. The cells were evaluated by flow cytometry to identify the changes in the cell cycle. The cell proliferation index was calculated according to the following formula: PI%=(S+G2/M)/ (G0/G1+S+G2/M). With the fourth sample, cells were loaded with 10 μmol/L 2′,7′-dichlorodihydrofluorescein for 30 min before harvesting. The ROS generation was assessed by fluorescence microscopy.
Detection of miR changes using microRNA chips in U266 cells after berberine treatment
Total miRs from cells (1×108) before and after treatment with 80 μmol/L berberine for 24 h were isolated using the mirVANATM miRNA Isolation Kit. The set of 186 miR genes was composed of 153 miRs identified in the miR registry at www.sanger.ac.uk/Software/Rfam, and 36 other miRs were manually selected from published papers or were found in the GenBank database at www.ncbi.nlm.nih.gov. Nineteen human miRs (10%) were found by homology with cloned miRs from other species (mainly mouse). For all of these miRs, we found the sequence of the precursor using the M Zucker RNA folding program at www.bioinfo.rpi.edu/applications/mfold/old/rna, and we selected the precursor sequence that gave the best score for the hairpin structure.
We used the Build 33 and Build 34 Version 1 of the Homo sapiens genome, which is available at www.ncbi.nlm.nih.gov/genome/guide/human. For each miR present in the database, we performed a BLAST search with the default parameters against the human genome to find the precise location followed by mapping using the maps available at the Human Genome Resources at the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov).
Detection of transcription factor activation by protein/DNA-binding arrays
The effect of berberine on multiple transcription factors in U266 cells was assessed using the TranSignal Protein/DNA Array from Panomics (Fremont, CA, USA). The results obtained by the protein/DNA array were further validated in DNA-binding assays for individual transcription factors by assaying nuclear extracts from berberine-treated or untreated cells using the Gel-Shift Kit from Panomics. All kits were used following the instructions of the manufacturer.
Luciferase reporter constructs and luciferase assay
The potential NF-κB-binding sites in the miR-21 promoter elements was predicted based on TFSEARCH (http://www.cbrc.jp/research/db/TFSEARCH.html) and MOTIF (http://motif.genome.jp/) database searches. miR-21 promoters were amplified by PCR from human genomic DNA with the following primers: sense, 5′-CCGCTCGAGTTTTGTTTTGCTTGGGAGGA-3′; and antisense, 5′-ATAAGAATGCGGCCGCAGCAGACAGTCAGGCAGGAT-3′. The PCR products were detected by agarose gel electrophoresis, and the DNA fragments were then isolated and cloned into the restriction enzyme-digested pmiR-CHECK™-2 Vector (Promega) using T4 DNA ligase (Fisher Scientific). All constructs were confirmed by sequencing. Mutations were introduced into the NF-κB-binding sites using the MagExtractor-Genome Kit (TOYOBO, Osaka, Japan), and cells were transfected with each reporter construct for 24 h. Luciferase activities were then measured using the Dual-Luciferase® Reporter Assay System (Promega, Madison, USA). The luciferase activity of each construct was compared with that of the promoterless pmiR-CHECK™-2 basic vector.
Assessing the DNA-binding activity of NF-κB by electrophoretic mobility shift assay (EMSA)
U266 cells were treated with or without berberine, washed twice with PBS, scraped in 1 mL HEGD buffer [25 mmol/L HEPES, 1.5 mmol/L EDTA, 1 mmol/L dithiothreitol, and 10% (v/v) glycerol (pH 7.6)], and homogenized. The cellular homogenate was centrifuged for 5 min at 14 000×g. The supernatant was discarded, and the pellet was suspended in 200 μL of HEGDK [25 mmol/L HEPES, 1.5 mmol/L EDTA, 1 mm dithiothreitol, 0.5 mol/L KCl, and 10% (v/v) glycerol (pH 7.6)] and incubated on ice for 1 h with frequent vortexing. Samples were centrifuged at 14 000×g for 1 min, and the nuclear protein concentration in the supernatants was determined. Proteins extracted from nuclei were incubated with biotin-labeled oligonucleotide probes corresponding to the miR-21 promoter region that contains the NF-κB-binding site (5′-CCACTCCTTTGTTAATACCAAGG-3′ and 5′-GAACATGATTGCTGAAAGAGTATGT-3′). The NF-κB competition control study was conducted using an unlabeled NF-κB consensus oligonucleotide. The samples were loaded and ran for 1 h at 30 mA.
Statistical analysis
All experiments were performed at least three times. Statistical analysis was performed with analysis of variance (ANOVA), followed by Turkey's t-test. P-values of <0.05 were considered statistically significant.
Results
Effects of berberine on U266 cell viability and IL-6 secretion
The viability of U266 cells was unaffected by treatment with 40 μmol/L berberine. However, after treatment with a high concentration of berberine (≥80 μmol/L) for more than 48 h, the growth of U266 cells was significantly reduced compared to that of the control group (P<0.05). U266 cells were affected by berberine treatment in a dose- and time-dependent manner (Figure 1A). When U266 cells were treated with berberine at different concentrations (40, 80, 120, and 160 μmol/L), the secretion of IL-6 was reduced by 9.6%, 27.8%, 53.7%, and 75%, respectively, at 24 h (Figure 1B).

Berberine inhibits U266 multiple myeloma cell viability and IL-6 secretion. (A) shows the dose- and time-dependent effect of berberine on U266 cell viability. (B) shows the secretion level of IL-6. Means±SD. n=3. bP<0.05 vs control.
Berberine treatment induces apoptosis in U266 cells
In each test group, berberine treatment increased the level of ROS and induced apoptosis and death in U266 cells (Figure 2A–2C). As shown in Figure 2D, when U266 cells were treated with 40, 80, 120, and 160 μmol/L berberine for 24 h, the number of cells in S phase was reduced, and a significantly higher number of cells were in the G2/M phase compared with the untreated control cells (P<0.001). Figure 2E shows the ultramicrostructure change in U266 cells after 80 μmol/L berberine treatment for 24 h. The morphological characteristics of normal U266 cells include the uniform distribution of cytoplasm with an integral cellular membrane (Figure 2Ea), and they have converted microvilli on the surface (Figure 2Ec). U266 cells treated with 80 μmol/L berberine exhibited morphological characteristics of apoptosis, including nuclear condensation, oversized cytoplasmic particles, oversized vacuoles, intact organelles and smooth, integral cellular membranes (Figure 2Eb). The microvilli disappeared, and ubiquitous emboli were observed (Figure 2Ed).

The effects of berberine on U266 cell cycle, apoptosis and ROS level. (A) FCM detected Annexin V and PI positive cells. (B)U266 cell apoptosis analysis using Hoechst 33342/PI stain. (C) U266 cell ROS analysis using DCFH-DA stain. (D) U266 cell cycle analysis. (E) U266 cell ultramicrostructure as detected by TEM and SEM. (Ea) Tumor cells were pleomorphic and of various sizes; their nuclei were large and unsymmetrical, with clear nuclear membranes, and were rich in euchromatin; and the cells contained chromatin particles, which were large intranuclear pseudo-inclusions. There were multiple visible nucleoli (sham group was stained with lead; ×3500). (Eb) Tumor cells were reduced in size and had karyopyknosis with chromatin margination and intercellular space widening (cells was stained with lead; ×5000). Chondriosome swelling and high electron dense apoptotic bodies were present, and vacuoles were present in the cytoplasm. (Ec) Morphological analysis of normal U266 cells (×5000) showed conferted microvilli. (Ed) After berberine treatment, invaginated microvilli and emboli were present. Means±SD. n=3. bP<0.05 vs control.
Berberine interferes with the expression of miR-21 in U266 cells and regulates a caspase-dependent apoptosis pathway
Based on the observed effect of berberine treatment on cell proliferation and viability, we selected the 80 μmol/L berberine condition for further mechanistic studies on miRs changes. According to the miR array, 56 miRs were expressed in the control group, and 64 miRs were expressed in the test group. Moreover, these data suggested that miR-21, which is closely associated with apoptosis, was significantly downregulated in test group. We used qRT-PCR to verify miR-21 levels after treatment with berberine (40, 80, 120, and 160 μmol/L) for 24 h, and we observed a clear trend of berberine dose-dependent downregulation of miR-21 (Figure 3A).

Berberine activates the mitochondrial apoptosis pathway and inhibits miR-21 expression in U266 cells. Expression of miR-21, Bcl-2, Bax, PUMA, caspase-3, and caspase-9 was detected in U266 cells treated with berberine (40, 80, 120, and 160 μmol/L) for 24 h (A and B). As compared to the control group, berberine-treated U266 cells had decreased levels of miR-21 and Bcl-2 and no change in Bax levels, but PUMA, caspase-3, and caspase-9 levels were all increased in a dose-dependent manner (B). We compared the miR-21 level in U266 between the cell transfected with miR-21 and treated with 80 μmol/L berberine (C). After the level of miR-21 was increased, the cell viability and IL-6 secretion increased (D and E), but the apoptosis ratio decreased (F and G). The MO and NC group are both positive control. The MO group contained U266 cells treated only with Lipofectamine 2000. The NC group was transfected with miR-NC, which had the same amount of basic groups but a different sequence. The MO, NC, and miR-21 were all chemically synthesized by Jima Com (Shanghai, China).
Bcl-2 family proteins play a critical role in the regulation of apoptosis by functioning as promoters (Bax) or inhibitors (Bcl-2). Bcl-2 is the target gene of miR-21. Based on the miR array, cell cycle analysis and apoptosis results, we measured the expression of apoptosis-related proteins, including Bcl-2, Bax, caspase-3, caspase-9, and PUMA, after treatment with 40, 80, 120, and 160 μmol/L berberine for 24 h. Berberine decreased the expression of Bcl-2 and increased the expression of PUMA, caspase-3, and caspase-9, but berberine did not change the expression levels of Bax (Figure 3B).
To verify that miR-21 was a key factor in the effects of berberine on U266 cells, we overexpressed miR21 by approximately 10-fold compared to the control group and by 20-fold compared to the berberine group using a transfection mimic of miR21 (Figure 3C). In accordance with the above mentioned findings, we used 80 μmol/L berberine as the effective concentration for the reverse test. MTT, ELISA, FCM, and morphology assays were performed to determine U266 cell responses to miR-21 and berberine. These data indicated that the upregulation of miR-21 expression promotes U266 cell proliferation and secretion of IL6 counteracting the effect of berberine (Figure 3D–3G).
Database analysis of transcription factor activation and potential binding sites in miR-21 promoter elements in U266 cells
MicroRNAs exhibit tissue-specific or developmental stage-specific expression and respond to extracellular stimuli, thereby indicating that their expression is tightly controlled. Most human miRNA genes, including miR-21, are transcribed by polymerase II (pol II) and can be classified as class II genes, in addition to all protein-coding genes. Human miRs are elaborately controlled through various regulatory mechanisms, including transactivation and transrepression, by nuclear transcription factors. We prepared nuclear extracts from berberine-treated and untreated U266 cells, and we compared the DNA-binding capacity of several transcription factors in the two nuclear extract samples. We found modulation of the binding activity of several transcription factors. However, the most consistent and notable results included an approximate 50% downregulation of the DNA-binding activity of NF-κB following treatment with berberine (Figure 4A). After inhibiting NF-κB by siRNA, the miR-21 level significantly decreased (P<0.01; Figure 4B). Binding of NF-κB subunits to the promoter elements of miR-21 has been implicated in the transcriptional regulation of miR-21. Two potential binding sites in the putative promoter element of the mir-21 gene are GGGAATTTTC (+1167 to +1176) and GGGAATTCTC (+1395 to +1404)12. The EMSA revealed decreased binding of p65 to the binding site at +1167 and +1395 of the mir-21 promoter element in berberine-treated cells (Figure 4C), and this repression was reversed by a single base mutation in the binding site (miR-21-mt) (Figure 4D). These results suggested that the complementary site in the miR-21 3′-UTR is a direct target of NF-κB-mediated posttranscriptional gene silencing.

Berberine inhibits intranuclear NF-κB activity and subsequently decreases transcriptional regulation of miR-21. The activity of promoter elements in the nucleoprotein was detected after treatment with or without 80 μmol/L berberine for 24 h. The four dots in the red circle were used to analyze repeated measurement data of NF-κB in a single sample (A). Berberine decreased NF-κB levels at the mRNA level (B). The NF-κB DNA-binding activity was inhibited with a C to G mutation in the promoter region of miR-21 (GGGAATTCTC; +1395 to +1404). The inhibition was confirmed by EMSA and luciferase assays (C and D). Mean±SD. n=3. bP<0.05 vs control.
Berberine upregulates lysine methyltransferase Set9
To investigate how berberine interferes with the effect of NF-κB, we performed a mRNA chip of U266 cells with or without 80 μmol/L berberine treatment for 24 h, and the results indicated that Set9 was highly expressed in U266 cells after berberine treatment (data not shown). We measured Set9 mRNA and protein levels after treatment with berberine at different concentrations for 24 h using qRT-PCR and Western blot, respectively (Figure 5A and 5B), and the results indicated that berberine increased the level of Set9 in a dose-dependent manner. Methylation of non-histone proteins is important for regulating their functions. Methylation of RelA inhibits NF-κB action by inducing the proteasome-mediated degradation of promoter-associated RelA. RelA is restricted to the nucleus, and this modification also regulates the promoter binding of p6513,14. We verified this phenomenon by knocking down Set9. The following two sets of human Set9 siRNAs significantly suppressed Set9 in U266 cells at both mRNA and protein levels: siRNA1, 5′-GCCUUGUAGGAGAAGUAAAtt-3′ (sense) and 5′-UUUACUUCUCCUACAAGGCtt-3′ (antisense); and siRNA2, 5′-GGGUUUAUGUUGCUGAAUCtt-3′ (sense) and 5′-GAUUCAGCAACAUAAACCCtt-3′ (anti-sense) (Figure 5C and 5D). Silencing Set9 by siRNA was partly due to the turnover level of NF-κB in the nucleoprotein (Figure 5D). The inhibitory effect of berberine on U266 cell proliferation was also partly counteracted by Set9-siRNA (Figure 5E).

Berberine upregulates lysine methyltransferase Set9 resulting in methylation of RelA and induction of proteasome-mediated degradation of NF-κB. The mRNA and protein levels of Set9 were measured in U266 cells treated with different concentrations of berberine for 24 h (A and B). (C) shows the mRNA level of Set9 by QRT-PCR method. After knocking down the Set9 by siRNAs the RelA level in nuclear was increased(D). And the cell viability was increased (E). Mean±SD. n=3. bP<0.05 vs control and eP<0.05 vs berberine group.
Discussion
The study of ancient herbal medicines may lead to novel cancer treatment strategies. berberine is a naturally occurring isoquinoline alkaloid that has anti-tumor effects on many types of cancer. Iizuka15 showed that berberine has an anti-cachectic effect in nude mice bearing human esophageal cancer cells and colon 26/clone 20 carcinoma cells and that this effect is mediated by an IL-6-related mechanism16. For tumor-bearing patients treated with berberine, nutritional parameters (eg, serum glucose level, wasting of adipose tissue and wasting of muscle tissue) change slightly, but IL-6 levels significantly decreased. Approximately 70% of multiple myeloma patients are over 60 years old, and 90% of multiple myeloma patients are over 50 years old. Almost all advanced stage patients are cachectic. The Karnofsky score and nutritional parameters help to indicate whether patients will be able to tolerate chemotherapy. IL-6 is an inflammatory cytokine that is closely related to nutritional metabolism, and it is also the key growth factor for multiple myeloma cells. Thus, we hypothesized that berberine may be an effective chemotherapeutic agent against multiple myeloma17,18. We found that berberine (80–160 μmol/L) was markedly cytotoxic to U266 cells and that it reduced the secretion of IL-6 in a dose- and time-dependent manner. We examined the ultramicrostructure of U266 cells by TEM and found karyopyknosis, chromatin margination, and cytoplasmic vacuoles, as well as other signs of apoptosis. Moreover, using SEM, we found that the microvilli on the surface of U266 cells almost completely disappeared after berberine treatment. Perhaps further research will help us to know whether berberine would induces multiple myeloma differentiation too. The present results suggested that IL-6 secretion may be influenced by cell apoptosis. The statistical difference between the control and 40 μmol/L berberine-treated groups at 24 h verified that small doses of berberine can suppress IL-6 secretion. Our results indicated that the viability of U266 cells significantly decreased after berberine treatment for 48 and 72 h, which is in contrast to a recent report by Li and colleagues19, who demonstrated that berberine treatment for 24 h upregulates the multidrug-resistant transporter (Pgp-170) in two oral (KB and OC2), two gastric (SC-M1 and NUGC-3) and two colon (COLO 205 and CT 26) cancer cell lines. Li and colleagues showed that pretreatment of cells with 32 μmol/L berberine for 24 h prior to paclitaxel treatment enhances cell viability and blocks the paclitaxel-induced G2/M arrest and morphological changes. Thus, they suggested that berberine modulates the expression and function of Pgp-170 and causes reduced responses to paclitaxel in digestive track cancer cells19. In the present study, we also measured the level of Pgp-170 by FCM, but we did not notice any significant changes in Pgp-170 levels in multiple myeloma cells after berberine treatment. Similar to the observation by Lin20, we found that berberine arrested U266 cells in the G2/M phase of the cell cycle in a dose-dependent manner. Thus, the effects of berberine on different cancer cells may not be comparable.
Apoptosis is known to be tightly regulated by anti-apoptotic and pro-apoptotic effector molecules21. In the present study, berberine also increased intracellular ROS in a dose-dependent manner. Many naturally occurring compounds have been reported to exert anti-cancer effects through ROS induction. For example, saikosaponin-a (SSa) and saikosaponin-d (SSd), which are two major pharmaceutical ingredients in Bupleurum radix, augment the cytotoxic effects of cisplatin through the induction of O2− and H2O2 accumulation in hepatic, pancreatic, breast and lung cancer cells. Pretreatment of cells with ROS scavengers effectively inhibited the potentiated cytotoxicity22. One possibility is that berberine induced cell apoptosis through inducing the generation of intracellular ROS. Early studies have suggested that ROS accumulation in cells results in pro-apoptotic Bcl-2 family protein activation23. miRNA profiling studies have shown that miR-21 is frequently overexpressed in many tumors and affects tumor progression by downregulating various target tumor suppressor genes, such as phosphatase and tensin homolog deleted on chromosome 10 (PTEN), PDCD4, and Bcl-224,25,26,27. We measured the levels of three hundred miRs in the U266 myeloma cell line with or without berberine treatment by miR array and found that the level of miR-21 significantly decreased with berberine treatment. We also studied the contribution of different Bcl-2 family proteins, including Bcl-2 and Bax, to elucidate the mechanism of berberine-induced apoptosis of U266 cells. There were no significant changes in the level of Bax, but there was a significant reduction in Bcl-2 levels. An increase in the Bax/Bcl-2 ratio stimulates the release of cytochrome c from mitochondria into the cytosol, which activates caspase-3 and caspase-9. The present results confirmed the concomitant execution phase of apoptosis. To confirm that miR-21 takes part in the effect of berberine on U266 cells, we induced the expression of miR-21 prior to berberine treatment and found that the apoptosis signs were significantly but incompletely blocked. There are two possible explanations for this observation as follows: 1) one gene may be regulated by many miRs, and miR-21 may be only one of many key factors; and 2) Lipofectamine 2000 may be cytotoxic to U266 cells, which may partly counteract the anti-apoptotic effect of miR-21.
Most research efforts on miRNAs have been focused on their physiological and pathological functions, and the molecular mechanisms of transcriptional regulation of miRNA genes remain largely unexplored. The majority of miRNA genes are located at intergenic regions or in antisense orientation to annotated genes, thereby suggesting that they may form independent transcription units. Other miRNAs are found in intronic regions, which may be transcribed as part of the annotated genes. Many oncogenes and chemopreventive chemicals can activate NF-κB in tumorigenesis and tumor progression28,29. A recent study has shown that p65 activates the miR-21 gene promoter and increases miR-21 RNA levels30. Although miR-21 is an NF-κB-transactivating gene, miR-21 works within the inflammation/transformation positive feedback loop by downregulating PTEN expression to increase NF-κB activity31,32. Through EMSA and luciferase analyses, we also demonstrated that NF-κB directly regulated the transcription of miR-21.
Constitutively active NF-κB is frequently encountered in a wide variety of tumors and in inflammation. Cells use multiple mechanisms for the decreasing of NF-κB activity, including NF-κB-dependent resynthesis of IκBα and the deubiquitination of upstream signaling molecules by deubiquitinating enzymes33. Recent studies have also indicated that proteasome-mediated degradation of nuclear DNA-bound NF-κB provides another layer of termination independent of negative feedback regulation. Set9 (also known as Set7)-mediated lysine methylation has recently emerged as a key posttranslational modification that regulates the function of histone and non-histone proteins, including NF-κB. RelA has been confirmed to be monomethylated by Set9 at lysine residues 314 and 315 both in vitro and in vivo, by using mutational and mass spectrometric analyses34,35,36,37. Methylation of RelA induces proteasome-mediated degradation of promoter-associated RelA, which inhibits NF-κB action38,39. The present study showed that Set9 increased in a berberine dose-dependent manner. Proteasome-mediated degradation of promoter-bound NF-κB is critical for the postinduction inactivation of nuclear NF-κB independent of IκBα resynthesis. Twelve hours after the transfection of U266 cells with siRNAs targeting Set9, the effects of berberine were partly offset by the recovery of NF-κB activity in nucleoproteins. In addition to methylation, RelA is also subject to a variety of posttranslational modifications, including acetylation and phosphorylation. Thus, we suggest that Set9 is not the only factor that suppresses NF-κB activity.
In conclusion, the present study demonstrated that berberine inhibits the proliferation of human multiple myeloma cells, as well as increases the intracellular ROS level, induces G2/M phase arrest and induces apoptosis. It is plausible that berberine negatively regulates NF-κB through Set9-mediated lysine methylation, which leads to a decrease in miR-21 levels and, in turn, an decrease in Bcl-2 levels, thereby triggering apoptosis.
Author contribution
Prof Hong-bo GUO and Mei-xia ZHANG designed the research and revised the manuscript; Hai-yan HU conducted the research; Xiu-ju WANG wrote the first draft and analyzed all the FCM data; Kun-peng LI did the statistic analysis, Yuan LIU, Zhi-gang LU, and Rui-hong DONG helped with portions of the research.
References
Katanoda K, Saika K . Comparison of time trends in multiple myeloma mortality (1990–2006) between countries based on the WHO mortality database. Jpn J Clin Oncol 2011; 41: 444–5.
Yin J, Zhang H, Ye J . Traditional chinese medicine in treatment of metabolic syndrome. Endocr Metab Immune Disord Drug Targets 2008; 8: 99–111.
Kuo CL, Chi CW, Liu TY . The anti-inflammatory potential of berberine in vitro and in vivo. Cancer Lett 2004; 203: 127–37.
Lin JP, Yang JS, Wu CC, Lin SS, Hsieh WT, Lin ML, et al. Berberine induced down-regulation of matrix metalloproteinase-1, -2 and -9 in human gastric cancer cells (SNU-5) in vitro. In Vivo 2008; 22: 223–30.
Tang J, Feng Y, Tsao S, Wang N, Curtain R, Wang Y . Berberine and Coptidis rhizoma as novel antineoplastic agents: a review of traditional use and biomedical investigations. J Ethnopharmacol 2009; 126: 5–17
Yount G, Qian Y, Moore D, Basila D, West J, Aldape K, et al. Berberine sensitizes human glioma cells, but not normal glial cells, to ionizing radiation in vitro. J Exp Therapeutics Oncol 2004; 4: 137–43.
Lin CC, Kao ST, Chen GW, Chung JG . Berberine decreased N-acetylation of 2-aminofluorene through inhibition of N-acetyltransferase gene expression in human leukemia HL-60 cells. Anticancer Res 2005; 25: 4149–55.
Yu FS, Yang JS, Lin HJ, Yu CS, Tan TW, Lin YT, et al. Berberine inhibits WEHI-3 leukemia cells in vivo. In Vivo 2007; 21: 407–12.
Lin CC, Kao ST, Chen GW, Ho HC, Chung JG . Apoptosis of human leukemia HL-60 cells and murine leukemia WEHI-3 cells induced by berberine through the activation of caspase-3. Anticancer Res 2006; 26: 227–42.
Wu SN, Yu HS, Jan CR, Li HF, Yu CL . Inhibitory effects of berberine on voltage- and calcium-activated potassium currents in human myeloma cells. Life Sci 1998; 62: 2283–94
Kim S, Kim Y, Kim JE, Cho KH, Chung JH . Berberine inhibits TPA-induced MMP-9 and IL-6 expression in normal human keratinocytes. Phytomedicine 2008; 15: 340–7.
Shin VY, Jin H, Ng EK, Cheng AS, Chong WW, Wong CY, et al. NF-PE targets miR-16 and miR-21 in gastric cancer: involvement of prostaglandin E receptors. Carcinogenesis 2011; 32: 240–5.
Liu Q, Jiang H, Liu Z, Wang Y, Zhao M, Hao C, et al. Berberine radiosensitizes human esophageal cancer cells by downregulating homologous recombination repair protein RAD51. PLoS One 2011; 6: e23427.
Choi YS, Park JA, Kim J, Rho SS, Park H, Kim YM, et al. Nuclear IL-33 is a transcriptional regulator of NF-κB p65 and induces endothelial cell activation. Biochem Biophys Res Commun 2012; 421: 305–11.
Iizuka N, Hazama S, Yoshimura K, Yoshino S, Tangoku A, Miyamoto K, et al. Anticachectic effects of the natural herb Rhizoma Coptidis and berberine on mice bearing colon 26/clone 20 adenocarcinoma. Int J Cancer 2002; 99: 286.
Iizuka N, Miyamoto K, Hazama S, Yoshino S, Yoshimura K, Okita K, et al. Anticachectice ffects of Rhizoma Coptidis, ananti-inflammatory herb, ones ophageal cancer cells that produce interleukin 6. Cancer Lett 2000; 158: 35.
Zipori D . The hemopoietic stem cell niche versus the microenvironment of the multiple myeloma-tumor initiating cell. Cancer Microenviron 2010; 3: 15–28.
Xie JY, Li MX, Xiang DB, Mou JH, Qing Y, Zeng LL, et al. Elevated expression of APE1/Ref-1 and its regulation on IL-6 and IL-8 in bone marrow stromal cells of multiple myeloma. Clin Lymphoma Myeloma Leuk 2010; 10: 385–93.
Li B, Fu J, Chen P, Zhuang W . Impairment in immunomodulatory function of mesenchymal stem cells from multiple myeloma patients. Arch Med Res 2010; 41: 623–33.
Lin SS, Chung JG, Lin JP, Chuang JY, Chang WC, Wu JY, et al. BBR inhibits arylamine N-acetyltransferase activity and gene expression in mouse leukemia L 1210 cells. Phytomedicine 2005; 12: 351–8.
Weyhenmeyer B, Murphy AC, Prehn JH, Murphy BM . Targeting the anti-apoptotic bcl-2 family members for the treatment of cancer. Exp Oncol 2012; 34: 192–9.
Wang Q, Zheng XL, Yang L, Shi F, Gao LB, Zhong YJ, et al. Reactive oxygen species-mediated apoptosis contributes to chemosensitization effect of saikosaponins on cisplatin-induced cytotoxicity in cancer cells. J Exp Clin Cancer Res 2010; 29: 159.
Feugang JM, Ye F, Zhang DY, Yu Y, Zhong M, Zhang S, et al. Cactus pear extracts induce reactive oxygen species production and apoptosis in ovarian cancer cells. Nutr Cancer 2010; 62: 692–9.
Lou Y, Yang X, Wang F, Cui Z, Huang Y . MicroRNA-21 promotes the cell proliferation, invasion and migration abilities in ovarian epithelial carcinomas through inhibiting the expression of PTEN protein. Int J Mol Med 2010; 26: 819–27.
Sayed D, He M, Hong C, Gao S, Rane S, Yang Z, et al. MicroRNA-21 is a downstream effector of AKT that mediates its antiapoptotic effects via suppression of Fas ligand. J Biol Chem 2010; 285: 20281–90.
Bhatti I, Lee A, James V, Hall RI, Lund JN, Tufarelli C, et al. Knockdown of microRNA-21 inhibits proliferation and increases cell death by targeting programmed cell death 4 (PDCD4) in pancreatic ductal adenocarcinoma. J Gastrointest Surg 2011; 15: 199–208.
Shi L, Chen J, Yang J, Pan T, Zhang S, Wang Z, et al. MiR-21 protected human glioblastoma U87MG cells from chemotherapeutic drug temozolomide induced apoptosis by decreasing Bax/Bcl-2 ratio and caspase-3 activity. Brain Res 2010; 1352: 255–64.
Ma X, Becker Buscaglia LE, Barker JR, Li Y . MicroRNAs in NF–kappaB signaling. J Mol Cell Biol 2011; 3: 159–66.
Zhou R, Hu G, Gong AY, Chen XM . Binding of NF-kappaB p65 subunit to the promoter elements is involved in LPS-induced transactivation of miRNA genes in human biliary epithelial cells. Nucleic Acids Res 2010; 38: 3222–32.
Niu J, Shi Y, Tan G, Yang CH, Fan M, Pfeffer LM, et al. DNA damage induces NF-κB-dependent microRNA-21 up-regulation and promotes breast cancer cell invasion. J Biol Chem 2012; 287: 21783–95.
Ruan Q, Wang T, Kameswaran V, Wei Q, Johnson DS, Matschinsky F, et al. The microRNA-21-PDCD4 axis prevents type 1 diabetes by blocking pancreatic beta cell death. Proc Natl Acad Sci U S A 2011; 108: 12030–5.
Marquez RT, Wendlandt E, Galle CS, Keck K, McCaffrey AP . MicroRNA-21 is upregulated during the proliferative phase of liver regeneration, targets Pellino-1, and inhibits NF-kappaB signaling. Am J Physiol Gastrointest Liver Physiol 2010; 298: G535–41.
Hayden MS, Ghosh S . Shared principles in NF-κB signaling. Cell 2008; 132: 344–62.
Ko S, Ahn J, Song CS, Kim S, Knapczyk-Stwora K, Chatterjee B . Lysine methylation and functional modulation of androgen receptor by Set9 methyltransferase. Mol Endocrinol 2011; 25: 433–44.
Yang J, Huang J, Dasgupta M, Sears N, Miyagi M, Wang B, et al. Reversible methylation of promoter-bound STAT3 by histone-modifying enzymes. Proc Natl Acad Sci U S A 2010; 107: 21499–504.
Gaughan L, Stockley J, Wang N, McCracken SR, Treumann A, Armstrong K, et al. Regulation of the androgen receptor by SET9-mediated methylation. Nucleic Acids Res 2011; 39: 1266–79.
Yang XD, Tajkhorshid E, Chen LF . Functional interplay between acetylation and methylation of the RelA subunit of NF-kappaB. Mol Cell Biol 2010; 30: 2170–80.
Ea CK, Baltimore D . Regulation of NF-kappaB activity through lysine monomethylation of p65. Proc Natl Acad Sci U S A 2009; 106: 18972–7.
Yang XD, Huang B, Li M, Lamb A, Kelleher NL, Chen LF . Negative regulation of NF-kappaB action by Set9-mediated lysine methylation of the RelA subunit. EMBO J 2009; 28: 1055–66.
Acknowledgements
This paper was supported by Guangdong Science and Technology Plan (No 2012A030400012).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Hu, Hy., Li, Kp., Wang, Xj. et al. Set9, NF-κB, and microRNA-21 mediate berberine-induced apoptosis of human multiple myeloma cells. Acta Pharmacol Sin 34, 157–166 (2013). https://doi.org/10.1038/aps.2012.161
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2012.161
Keywords
This article is cited by
-
Advanced application of nanotechnology in active constituents of Traditional Chinese Medicines
Journal of Nanobiotechnology (2023)
-
Tumor-suppressive functions of protein lysine methyltransferases
Experimental & Molecular Medicine (2023)
-
Discovery of the oncogenic MDM2, a direct binding target of berberine and a potential therapeutic, in multiple myeloma
Functional & Integrative Genomics (2022)
-
Berberine reduces temozolomide resistance by inducing autophagy via the ERK1/2 signaling pathway in glioblastoma
Cancer Cell International (2020)
