
Abstract
This study examined the genetic contribution of single male immigrants to small, inbred laboratory populations of Drosophila melanogaster. Genetic contribution was assessed by measuring the relative frequency of immigrant marker alleles in the first and second generations after immigration, while controlling for any selection effects at the marker locus, and for the experience of male immigrants. When immigrants were outbred, the mean frequency of the immigrant allele was significantly higher than its initial frequency, in both the first and second generations after immigration. There was no significant change in allele frequency for populations receiving inbred immigrants. The increase in allele frequency for outbred immigrants was attributed to an initial outbred vigour fitness advantage of immigrant males over resident males experiencing inbreeding depression. Hybrid vigour of immigrant progeny and the rare-male effect did not have a statistically significant role in the fitness advantage of the immigrant allele. The results suggest that inbreeding may have a considerable impact on the contribution of immigrants to the genetic diversity of populations.
Similar content being viewed by others

Introduction
Wright’s infinite-island model (Wright, 1931) has been widely used as the basis for estimating immigration rates within subdivided populations. Such widespread use of this model recently received criticism on the grounds that many real systems do not meet its assumptions (Whitlock & McCauley, 1999). Whitlock & McCauley (1999) argued that Wright’s model is unlikely to produce reliable estimates of the true rate at which immigrant individuals arrive into populations, as compared to the effective genetic rate of immigration. Essentially this means that biologists may have been putting false hopes in using Wright’s model to link the genetic and ecological impacts of immigration.
Importantly, Wright’s model assumes that immigrants have the same fitness as population residents. For real systems however, there are several reasons why this assumption may be violated. For example immigrants may differ from population residents in both age and social status (Gaines & McClenaghan, 1980).
In this study we examined the effect of inbreeding on immigrant fitness. In particular we examined the genetic contribution of single male immigrants (both inbred and outbred) arriving into inbred populations. Essentially this addresses two assumptions implicit in Wright’s model: (i) that immigrants have the same fitness as residents of the populations they arrive into, and (ii) that an immigrant’s fitness is independent of whether it originated from a large outbred source population or a small inbred source population.
We initially identified three reasons why immigrants arriving into inbred populations might have higher fitness than residents. These were: (i) higher fitness over resident males due to a rare-male effect; (ii) higher fitness over resident males experiencing inbreeding depression (we refer to this as ‘initial outbred vigour’); and (iii) higher fitness caused by hybrid vigour of immigrant progeny over inbred competitors.
The rare-male hypothesis states that genetically rare males have greater mating success over their nonrare competitors due to rareness per se. Although the underlying mechanisms are poorly understood, it has been suggested for Drosophila that females assess the rarity of the different males in a population using a range of cues, and subsequently choose rare males to mate with them (Petit & Ehrman, 1969). This rare-male effect has been reported in a number of Drosophila species (Petit & Ehrman, 1969), as well as a number of other insect species (Sinnock, 1970; Grant et al., 1974). Rareness was expected to arise in the present study because of the random drift of allele frequencies whenever populations were inbred.
Inbreeding depression is the decrease in fitness resulting from the mating of closely related individuals. The prevailing belief is that this drop in fitness is caused by increased homozygosity associated with inbreeding, which in turn causes more frequent expression of rare recessive (or partially recessive) deleterious alleles (Charlesworth & Charlesworth, 1987; Lande, 1988). In the present study inbreeding had the potential to favour the immigrant genome at two points in time. First, it could give outbred immigrants an initial outbred vigour advantage over inbred residents. Secondly, it could give immigrant progeny a hybrid vigour advantage over inbred competitors. Hybrid vigour arises when the crossing of two inbred lines (or an outbred line with an inbred line) produces individuals that are heterozygous at loci that were previously homozygous for deleterious alleles (Falconer, 1981).
We studied the impact of these mechanisms by measuring the genetic contribution of single male immigrants arriving into inbred populations of Drosophila melanogaster in the laboratory. This species was chosen because of its fast generation time, the ease with which sexes can be distinguished, evidence that the species is highly polymorphic at allozyme loci (Singh & Rhomberg, 1987), and ease of rearing. Furthermore, indirect evidence suggested that single D. melanogaster immigrants are capable of making a substantial genetic contribution to small, inbred populations. Spielman & Frankham (1992) found that single D. melanogaster immigrants caused a marked decrease in the inbreeding depression of inbred laboratory populations. The present study differs from that of Spielman & Frankham (1992) by directly considering the implications of immigration on allele frequencies per se, rather than any effects on inbreeding depression.
Our experimental design involved two treatments: one where the immigrant was inbred and one where the immigrant was outbred. Inbred immigrants received the same inbreeding regime as the populations they arrived into. Such inbred immigrants may experience a fitness advantage over population residents because (i) they are genetically rare with respect to their recipient population, and (ii) their progeny experience a hybrid vigour advantage over the progeny of residents. Importantly however, there can be no initial outbred vigour in this case, as the immigrant males would, on average, have the same level of inbreeding depression as the resident males.
Outbred immigrants were much less inbred than their recipient populations. As with inbred immigrants, outbred immigrants may also be genetically rare with respect to their recipient population, and their offspring may also experience hybrid vigour. However, in addition outbred immigrants may themselves experience an initial outbred vigour advantage as a result of inbreeding depression amongst their recipient population. By comparing the two treatments we were therefore able to assess the importance of any initial outbred vigour.
Importantly, both a rare-male effect and hybrid vigour potentially give the immigrant genome a fitness advantage in the generations following immigration (compared with initial outbred vigour, which is restricted to the generation in which the immigrant arrives). Given this potential, we decided to measure the fate of the immigrant allele in both the first and the second generations after immigration.
The genetic contribution of single immigrants into populations has received some attention in the past. Kaufman & Wool (1992) used flour beetles to measure the frequency of a phenotypically dominant marker in the first generation after immigration. Although immigrant beetles had significantly higher fitness than residents, it is possible that this immigrant advantage was caused entirely by selection associated with the marker used. The use of a dominant marker restricts such an experiment to being performed in one direction, with homozygous dominant immigrants arriving into homozygous recessive populations. Importantly, we controlled for any effect of selection at the marker locus. By using a codominant allozyme polymorphism (Adh with two alleles F and S) we were able to perform the experiment reciprocally, producing one set of results with an FF immigrant arriving into an SS population, and another set vice versa. This reciprocal design is particularly important given that there is evidence of selection at the Adh locus in D. melanogaster (Oakeshott, 1979; Gilbert & Richmond, 1982). Secondly, we controlled for the experience of immigrants by rearing all experimental individuals (immigrant and resident) under equivalent conditions.
The structure of this study can be summarized as follows: (i) the fate of an immigrant allele was measured in the two generations following immigration; (ii) there were two main treatments: inbred immigrant and outbred immigrant; and (iii) the entire experiment was performed reciprocally, to account for any selection associated with the allelic marker used.
Methods
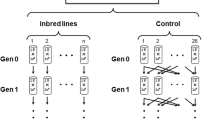
This study involved collecting a wild-type population from the field, developing two marked base populations (each fixed for a different allele at the same locus), and then using these marked populations as the source of individuals for the experiment. Figure 1 provides an overview of the methods that are detailed below.

Experimental design. This shows the processes leading from the initial collection of wild-type Drosophila melanogaster flies to the introduction of either an outbred SS or inbred SS immigrant into an inbred FF population.
Establishment and maintenance of wild-type laboratory population
A prerequisite for this experiment was to have a population which, when inbred, would have the potential to experience both a rare-male effect and inbreeding depression. This meant obtaining a base population that contained as much genetic diversity as possible. We therefore made a field collection of female D. melanogaster from a local winery and maintained the subsequent laboratory population as large as was practical. Twenty progeny from each of 90 wild-inseminated females were combined to create a total population of 1800 individuals. This was kept at approximately 2000 individuals in subsequent generations, with the culture maintained at 25°C as discrete generations, with approximately 30 days per generation. The flies were reared on an agar medium (containing cornmeal, treacle, baker’s yeast and methylparaben mould inhibitor) and were handled under anaesthesia using CO2.
Development of marked outbred base populations
Approximately 16 generations after the field collection, the wild-type laboratory population was used to set up two marked base populations — each one fixed for a different Adh allele (F and S). These base populations then became the source of all individuals (both inbred and outbred) in the experiments. Electrophoresis was carried out on cellulose acetate gels using methods described by Richardson et al. (1986). Although we found a number of polymorphic loci in the wild-type population, Adh was chosen because it had two common alleles and required the least expensive stain for large-scale screening. Although there is evidence of selection at this locus (Oakeshott, 1979; Gilbert & Richmond, 1982), we considered it better to use Adh than a poorly studied locus — in this way we would at least have some insights into any marker effect.
We founded the two marked base populations with as many individuals as possible in order to maximize their genetic diversity, and therefore maximize their potential to produce outbred and inbred individuals that would differ in fitness. The important issue here is that any outbred experimental fly could only be as outbred as the base population from which it was drawn. Therefore, a total of 140 wild-type virgin flies were nondestructively genotyped by removing a middle leg from individuals immobilized in a cold room (10°C), and squashing the leg directly onto the gel loading position. Individuals homozygous for the same allele were then arranged into mating pairs, which were placed in separate vials and allowed to reproduce. This provided a total of six SS pairs and six FF pairs. Each marked population was increased to 180 individuals in the following generation, and maintained at 2000 individuals in subsequent generations. The inbreeding imposed by this bottleneck is considered negligible relative to the two generations of full sib–sib mating imposed during experiments. If we set the inbreeding coefficient, F, to zero for the wild-type base population, then F=0.04 for outbred immigrants, and F=0.46 for both inbred residents and immigrants (based on Falconer (1981)). The purity of the two marked base populations was tested and confirmed both before and after individuals were removed for the experiments by sampling 20 individuals from each population.
The experiment
The units of replication in this study were single populations. Each replicate involved creating an inbred population founded by a mated female from one of the marked base populations. This is similar to founding each population with a mating pair of virgins given that D. melanogaster has a strong last-male advantage, with 85% of a female’s progeny being sired by her last mate (Gromko et al., 1984). Each population was then inbred for two generations by removing a single female every generation and placing her in a vial of fresh medium. Given the last-male advantage this approximates full sib–sib mating in each generation.
In the third generation (20 days after oviposition from the previous generation), 10 females and nine males were removed from the population, and placed on fresh medium together with a single male immigrant that was marked with the alternate allele to the resident flies. Importantly, immigrants were given similar experience to resident flies, whereby each immigrant was drawn from a vial in which a mated female had been allowed to oviposit 20 days beforehand. The mother of each outbred immigrant was drawn directly from the appropriate marked base population. In contrast, inbred immigrants had experienced the same inbreeding regime as the populations they arrived into. After the immigrant was introduced, each population was kept on fresh medium for 10 days to allow remating, and then placed in a jar with fresh medium for three days to allow females to oviposit.
When the immigrant arrived, the immigrant marker allele made up 1 in 20 (i.e. 0.05) of the total alleles in each replicate population. The null hypothesis was that the mean allele frequency would not change from this initial frequency over time. Allele frequency was measured by sampling up to 20 individuals in the two generations following immigration. Each generation was discrete, and was separated by 30–33 days. On day 20 of the first generation, 10 males and 10 females were removed from each population and placed on fresh medium (the remaining flies were discarded). Ten days later these 20 flies were placed on fresh medium and allowed to oviposit for three days. These same flies were then frozen and genotyped. This meant that the flies that sired the second generation were the same individuals genotyped in the first generation. Although less than 20 individuals were available for some replicates, very few (only 2.5% of all replicates) provided fewer than 15 individuals.
A total of 40 replicate populations were established with FF-marked immigrants arriving into SS populations. In 20 of these the immigrant was outbred and in the other 20 the immigrant was inbred. Because allele frequency was measured over two generations, this experimental design provided four data sets. Although these will be referred to as four separate ‘treatments’, the data sets from the two generations are essentially repeated measures rather than separate experimental treatments. A reciprocal experiment (with equal replication) was performed in which SS immigrants were introduced into FF populations. The entire experiment of eight treatments was carried out simultaneously. All replicates were independent in the sense that each replicate was derived from a separate female drawn from one of the marked base populations. The populations of all treatments were thoroughly shuffled to minimize bias caused by microclimatic variation in laboratory conditions.
Two sets of questions were addressed in the data analyses.
1 Was there a change in the mean allele frequency from the null level of 0.05 (one immigrant fly in a population of 20) for each of the treatments?
2 Were there differences in the mean allele frequencies between certain pairs of treatments (see results for details)?
All data were analysed using α=0.05 as the significance level, and all analyses were performed as two-tailed tests to include the possibility that immigrants could somehow be at a selective disadvantage in inbred populations. Importantly, the data were not pooled over markers. Our concern was that in the presence of a strong marker effect, pooling would produce an immigrant allele frequency greater than the null frequency of 0.05, and thereby give a false impression of an immigrant fitness advantage.
Results
Statistical tests
The frequency distributions of the immigrant allele frequency in the first and second generations after immigration were not normally distributed (Fig. 2). Furthermore, data transformation was of little value given the large numbers of zeros in these distributions. Nonetheless the use of parametric analyses was considered appropriate by virtue of the central limit theorem, which states that even if a distribution is highly skewed, the probability distribution of sample means will approximate a normal distribution as the number of replicates in the sample increases (Hays, 1988).

Distributions of immigrant allele frequency in Drosophila melanogaster. Shown for each treatment are the numbers of replicates (from n=20) belonging to different allele frequency categories. Each category has a width of 0.10, and each label on the x-axis refers to the midpoint of a category. The grey portion of the 0.05 category represents the number of values that were zeros. The sample mean and sample variance are given for each treatment.
Comparison of treatment means to the null allele frequency
The mean allele frequency was compared to the null allele frequency of 0.05 for each of the eight treatments. The test in each case was whether a 95% confidence interval around the sample mean would include the null allele frequency of 0.05. Although the sample mean was greater than the null allele frequency for all treatments, it was significantly greater only when the immigrant was outbred (Fig. 3). This was true for both the first and second generations after immigration, and for both marker combinations.

Sample mean (95% confidence interval) for all treatments. First and second refer to generations after immigration. Note the position of confidence intervals relative to the null allele frequency.
Comparison of allele frequency between treatment means
Treatment means were compared in a pair-wise manner using t-tests. The four types of comparisons were:
1 first generation (outbred vs. inbred immigrant);
2 second generation (outbred vs. inbred immigrant);
3 inbred immigrant (second generation vs. first generation);
4 outbred immigrant (second generation vs. first generation).
Although non-normality and heterogeneity of variances (Fig. 2) initially threw concerns on using t-tests, this was considered a robust approach for the data at hand. In regards to normality the central limit theorem once again applies (Sokal & Rohlf, 1981, p. 414), while the heterogeneity of variances is compensated by the equality of sample sizes — an important component of t-test robustness (Zar, 1984, p. 130).
Of the four t-tests performed on FF immigrant data, only first generation (outbred vs. inbred immigrant) showed a significant difference between treatment means (Table 1). Of the four tests applied to the SS immigrant data, two showed a significant difference between treatment means (Table 1). These were first generation (outbred vs. inbred immigrant), and second generation (outbred vs. inbred immigrant). Thus, unlike the SS immigrant data, the FF immigrant data did not show a significant difference for second generation (outbred vs. inbred immigrant). Notably however, this comparison was only marginally nonsignificant for FF immigrants, with P=0.064 (Table 1).
Results summary
The reciprocal marker experiments produced very similar results (Fig. 3). In summary:
1 When the immigrant was outbred, the mean frequency of the immigrant allele was significantly higher than the null allele frequency of 0.05 in the first and second generations after immigration.
2 When the immigrant was inbred, the mean frequency of the immigrant allele was not significantly different from the null allele frequency in either the first or second generations after immigration.
3 The mean allele frequency of both inbred and outbred immigrants of both markers did not change significantly from the first generation to the second generation.
4 In the first generation after immigration, the mean allele frequency of outbred immigrants was significantly higher than that of inbred immigrants.
5 In the second generation after immigration, the mean allele frequency of outbred immigrants was still significantly higher than that of inbred immigrants when the immigrant carried the SS marker. This difference was marginally nonsignificant for FF immigrants.
Discussion
This experiment showed that inbreeding can have a considerable impact on the contribution of single male immigrants to the genetic diversity of populations.
In addition, the rare-male effect and hybrid vigour of immigrant progeny seemed to have had little impact on immigrant fitness in this system. This is evident in the observation that the genetic contribution of inbred immigrants did not differ significantly from the initial null allele frequency of 0.05. However, although nonsignificant, the fact that the sample means for this treatment were above the null frequency for both markers suggests a trend worthy of further study.
The apparent lack of a rare-male effect in this study lends support to suggestions that this effect either (i) does not exist or (ii) is not of universal importance. In terms of the former, a number of authors have argued that the rare-male effect may be an artifact of experimental design (Bryant et al., 1980; Knoppien, 1987). In relation to the latter, it has been argued that the rare-male effect is not as strong for D. melanogaster as for another well studied species in this regard — Drosophila pseudoobscura (Markow et al., 1980).
The observation that inbred immigrants had no significant fitness advantage suggests that the rare-male effect and hybrid vigour were also negligible when the immigrant was outbred. This inference relies on the assumption that both the rare-male effect and hybrid vigour operate with equal strength irrespective of whether the immigrant is inbred or outbred. If this assumption is valid, the results imply that the fitness advantage of outbred immigrants over residents is attributable to initial outbred vigour, as this was the only mechanism identified as giving outbred immigrants an advantage over inbred immigrants. There are several ways in which this initial outbred vigour could occur. Outbred immigrants may have been (i) more active in courting mates; (ii) more attractive to females by appearing fitter than resident males (e.g. by being larger or more active); or (iii) more successful in fertilizing females (e.g. by producing semen that is more competitive than that of resident males). Exploration of the relative importance of these mechanisms provides opportunity for further study.
This experiment did not detect a significant change in the frequency of the immigrant allele from the first generation to the second generation after immigration. This was true for both markers and both inbred and outbred immigrants. This result is somewhat surprising given that first generation populations should include male immigrant progeny that, because of hybrid vigour, have the same advantage over their inbred competitors that gave the original male immigrants their initial outbred vigour advantage. One potential explanation is that the difference in fitness between immigrant progeny and inbred individuals was not as strong for females as it was for males. This would clearly reduce the average impact of hybrid vigour. Recombination of genes may also have reduced the impact of hybrid vigour, a mechanism that potentially applies to both sexes. The effect of recombination is that when immigrant progeny reproduced, the marker allele would have become disassociated from part of the immigrant genome that previously gave it a selective advantage.
Another issue to consider is that an increase in allele frequency between the first and second generations may have been obscured by increased variance. Indeed, in every situation (inbred and outbred immigrants, and both marker situations) the sample variance increased from the first to the second generation (Fig. 2). This is understandable when we consider the design of this experiment — from all treatments initially having the same immigrant allele frequency of 0.05, the variance in allele frequencies would tend to accumulate over the generations as population allele frequencies diverge. Although nonsignificant, there were consistent increases in the sample means from the first to second generations for all treatment combinations and we feel it is unwise to say that any increase can be ruled out. Therefore we suggest further testing of the ability of immigrant genes to increase in frequency over the generations following immigration.
Although this study controlled for both (i) a marker effect, and (ii) the experience of male immigrants relative to population residents, there remains the potential for inherited environmental effects to have influenced the results. Inherited environmental effects are those components of an individual’s phenotype that are derived from either parent, apart from nuclear genes (Rossiter, 1996). We have to acknowledge that the parents of outbred immigrants were reared in jars containing many hundreds of individuals, whereas the parents of all inbred immigrants were reared in small vials containing approximately 20–40 individuals. Although we believe the density of flies was comparable in each case, we cannot dismiss the possibility that inherited environmental effects influenced the results to some degree.
This study has demonstrated a situation where Wright’s (1931) assumptions of (i) immigrants having the same fitness as residents, and (ii) immigrants from different source populations having the same mean fitness as each other, are not met. Essentially this adds another element to our understanding of the complex relationship between the rate at which individuals immigrate into populations, and their impact in terms of gene flow. In doing this, our study lends support to the notion that population geneticists should be careful when using measures of population subdivision (e.g. FST) to infer Nm — the number of migrants entering a population per generation (Whitlock & McCauley, 1999).
The results of this study also have implications for conservation biology. It is thought that the occasional arrival of immigrants may rescue populations from extinction (Brown & Kodric-Brown, 1977), and that such a rescue effect may play a crucial role in the dynamics of metapopulations (Hanski et al., 1996). If genetic diversity contributes to such a rescue effect, our study suggests that even greater value should be placed on large mainland populations that act as a source of outbred immigrants to small inbred populations.
References
Brown, J. H. and Kodric-Brown, A. (1977). Turnover rates in insular biogeography: effect of immigration on extinction. Ecology, 58: 445–449.
Bryant, E. H., Kence, A. and Kimball, K. T. (1980). A rare-male advantage in the housefly induced by wing clipping and some general considerations for Drosophila. Genetics, 96: 975–993.
Charlesworth, D. and Charlesworth, B. (1987). Inbreeding depression and its evolutionary consequences. Ann Rev Ecol Syst, 18: 237–268.
Falconer, D. S. (1981). Introduction to Quantitative Genetics, 2nd edn. Longman Scientific and Technical, New York.
Gaines, M. S., McClenaghan, L. R., Jr. (1980). Dispersal in small mammals. Ann Rev Ecol Syst, 11: 163–196.
Gilbert, D. G. and Richmond, R. C. (1982). Studies of esterase 6 in Drosophila melanogaster XII. Evidence for temperature selection of Est 6 and Adh alleles. Genetica, 58: 109–119.
Grant, B., Snyder, G. A. and Glessner, S. F. (1974). Frequency-dependent mate selection in Mormoniella vitripennis. Evolution, 28: 259–264.
Gromko, M. H., Gilbert, D. G. and Richmond, R. C. (1984). In: Smith, R. L. (ed.) Sperm transfer and use in the multiple mating system of Drosophila, pp. 371–426, Academic Press, Orlando, FL.
Hanski, I., Moilanen, A. and Gyllenberg, M. (1996). Minimum viable metapopulation size. Am Nat, 147: 527–541.
Hays, W. L. (1988). Statistics, 4th edn. Holt, Rinehart and Winston Inc., New York.
Kaufman, B. and Wool, D. (1992). Gene flow by immigrants into isolated recipient populations: a laboratory model using flour beetles. Genetica, 85: 163–171.
Knoppien, P. (1987). Rare-male mating advantage: an artifact caused by differential storage conditions?. Behav Genet, 17: 409–425.
Lande, R. (1988). Genetics and demography in biological conservation. Science, 241: 1455–1460.
Markow, T. A., Richmond, R. C., Mueller, L., Sheer, I., Roman, S., Laetz, C. and Lorenz, L. (1980). Testing for rare male mating advantages among various Drosophila melanogaster genotypes. Genet Res, 35: 59–64.
Oakeshott, J. G. (1979). Selection affecting enzyme polymorphisms in laboratory populations of Drosophila melanogaster. Oecologia, 143: 341–354.
Petit, C. and Ehrman, L. (1969). Sexual selection in Drosophila. In: Dobzhansky, T., Hecht, M. K. and Steere, W. C. (eds) Evolutionary Biology, vol. 3, pp. 177–223. Appleton-Century-Crofts, New York.
Richardson, B. J., Baverstock, P. R. and Adams, M. (1986). Allozyme Electrophoresis: A Handbook for Animal Systematics and Population Studies. Academic Press, Sydney.
Rossiter, M. C. (1996). Incidence and consequences of inherited environmental effects. Ann Rev Ecol Syst, 27: 451–476.
Singh, R. S. and Rhomberg, L. R. (1987). A comprehensive study of genic variation in natural populations of Drosophila melanogaster II. Estimates of heterozygosity and patterns of geographic differentiation. Genetics, 117: 255–271.
Sinnock, P. (1970). Frequency dependence and mating behavior in Tribolium castaneum. Am Nat, 104: 469–476.
Sokal, R. R. and Rohlf, F. J. (1981). Biometry, 2nd edn. W. H. Freeman, San Francisco.
Spielman, D. and Frankham, R. (1992). Modeling problems in conservation genetics using captive Drosophila populations: improvement of reproductive fitness due to immigration of one individual into small partially inbred populations. Zoo Biol, 11: 343–351.
Whitlock, M. C. and McCauley, D. E. (1999). Indirect measures of gene flow and migration: FST≠ 1/(4Nm + 1). Heredity, 82: 117–125.
Wright, S. (1931). Evolution in Mendelian populations. Genetics, 16: 97–159.
Zar, J. H. (1984). Biostatistical Analysis, 2nd edn. Prentice Hall, Englewood Cliffs, NJ.
Acknowledgements
We thank Carolyn Horne, Terry Reardon and Soolim Carney for assistance with electrophoretic work, Sue Carthew for access to laboratory facilities, and Richard Frankham for advice on experimental design and rearing of Drosophila. We also value the constructive criticism of the manuscript by Jeanine Baker, and two reviewers.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ball, S., Adams, M., Possingham, H. et al. The genetic contribution of single male immigrants to small, inbred populations: a laboratory study using Drosophila melanogaster. Heredity 84, 677–684 (2000). https://doi.org/10.1046/j.1365-2540.2000.00721.x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1046/j.1365-2540.2000.00721.x
Keywords
This article is cited by
-
Genetic and fitness consequences of interpopulation mating in Dianthus guliae Janka: conservation implications for severely depleted and isolated plant populations
Conservation Genetics (2015)
-
Induced dispersal in wildlife management: experimental evaluation of the risk of hybrid breakdown and the benefit of hybrid vigor in the F1 generation
Conservation Genetics (2011)
-
Genetic rescue of an inbred captive population of the critically endangered Puerto Rican crested toad (Peltophryne lemur) by mixing lineages
Conservation Genetics (2010)
-
Gene Flow and Adaptive Potential in Drosophila melanogaster
Conservation Genetics (2006)
