
Abstract
Kit and its ligand stem cell factor (SCF) play a fundamental role in hematopoiesis, melanogenesis and gametogenesis. Homozygous Wv mutant mice with a mutation in kit show abnormalities in these cell lineages. Fas is a member of the death receptor family inducing apoptosis. In this study, we generated double-mutant mice (Wv/Wv:Fas−/−) and analyzed histologically their reproductive organs. In testes and ovaries of the double-mutant mice, testicular germ cells and oocytes were detected, respectively, whereas the same-aged Wv/Wv mice contained neither cells. In addition, inhibition of Kit signals by administration of anti-Kit mAb, which induces degeneration of testicular germ cells in vivo in wild-type mice, did not cause degeneration in Fas-deficient mice. In testicular germ cells of Wv/Wv mutant mice, an increase of Fas expression was observed in spermatogonia. Further, in vitro treatment with SCF was shown to downregulate Fas on fibroblasts expressing exogenous Kit through activation of PI3-kinase/Akt. All the results clearly indicate that Fas-mediated apoptosis is involved in germ cell degeneration accompanied by defects in Kit-mediated signals, and Kit signaling negatively regulates Fas-mediated apoptosis in vivo.
Similar content being viewed by others
Introduction
Kit is a receptor tyrosine kinase (RTK) belonging to the PDGF receptor/CSF-1 receptor family.1,2,3 Kit and its ligand, stem cell factor (SCF), play a fundamental role during the establishment and maintenance of the three stem cell populations: melanoblasts, hematopoietic stem cells and germ cells. Genes encoding Kit and SCF are mapped to the white spotting (W) and steel (Sl) loci in mice, respectively.4,5,6,7,8 Both mutant mice of W and Sl loci show similar phenotypes such as severe macrocytic anemia, a decreased number of mast cells, a complete lack of coat pigmentation and infertility.9 Further, many alleles of variable severity at both W and Sl loci have been identified.10 For example, Wv mutant mice with the C57BL/6 background have a point mutation in the cytoplasmic region of Kit, resulting in no coat pigmentation, severe anemia, a deficiency of mast cells and infertility in homozygotes (Wv/Wv).11,12
In reproductive organs, Kit plays essential roles in spermatogenesis and oogenesis.10 In the testes, Kit receptor expression is restricted to differentiating type A to type B spermatogonia, primary spermatocytes and Leydig cells.13,14,15,16 In particular, type A spermatogonia cells express a maximal amount of Kit at both the transcript and protein level, followed by type B spermatogonia.13,17,18 Therefore, the germ cells of the homozygous W mutant mice are impossible to develop and degenerate accompanied by apoptosis leading to infertility owing to a lack of Kit signals.18 In the ovaries, Kit is expressed on not only primordial and growing oocytes throughout the development of follicles but also on interstitial theca cells.13,18 In the ovaries of W mice, oogenesis completely fails because of defects of Kit signals resulting in no follicles and oocytes.10
Members of the RTK family play essential roles in the proliferation, differentiation, migration and survival of various cells.19 Kit as well as other RTKs interacts with several intracellular signal transducers including PI3-kinase, JAK2, STAT1, src-related kinases, SH2-containing tyrosine phosphatases and adaptor proteins such as SOCS1, Shc, GRB2 and GRB7. These signal transducers function in various cellular processes by activating target molecules including PKC, Akt and MAP kinases such as ERK and JNK.20 Kit is strongly suggested to be indispensable for not only a mitogenic response but also an antiapoptotic response.
Fas is a cell surface receptor belonging to the death receptor family, along with tumor necrosis factor receptor type1 (TNF-R1) and TRAIL receptors.21 Fas can induce cell death in a variety of tumors and lymphocytes by ligation with Fas ligand or an agonistic anti-Fas monoclonal antibody (mAb).22,23,24,25 While the physiological role of Fas has been well studied in the immune system,26,27,28,29 Fas mRNA is expressed at high levels in not only the thymus and spleen, but also the liver, heart, lungs and ovaries of mice.26,30 Therefore, it could be that Fas acts in the physiological deletion of potentially harmful or unnecessary cells in these tissues.28
In female gonads, we and others demonstrated that Fas is expressed on granulosa cells of various follicles and luteal cells.31,32,33,34,35,36,37,38 A few reports showed Fas expression on oocytes, but we have not been able to confirm this.32,38,39 The expression on granulosa and luteal cells suggests that Fas is involved in follicular atresia and luteolysis for the gonadal homeostasis of adult mice. In male gonads, Fas and Fas ligand are detected in the testes, but their function is still unclear. As Fas-deficient mice showed no abnormality in the testes,40 it was thought that Fas is not essential in germ cell homeostasis. However, recent reports insisted on the involvement of Fas in the physiological death of germ cells.41,42,43 In addition, Fas expression was shown to increase in the testes after drug treatment, ischemia–reperfusion or heat shock.44,45,46,47 In these damaged testes, Fas may play a role as a regulator of pathological germ cell death, although this has not been clearly determined.
To investigate the relation between Kit-mediated survival signals and Fas-mediated apoptotic signals in gonads, we generated double-mutant mice by intercrossing Kit-deficient Wv mutant mice with Fas knockout mice. Our study clearly demonstrated that a defect of Kit signaling leads to activation of a Fas-mediated apoptotic signal in both spermatogenesis and oogenesis, and Fas is involved in germ cell loss in Kit signaling-deficient mice. Moreover, our findings provide new evidence of crosstalk of signals via Kit and Fas, indicating that Kit-induced signals negatively regulate Fas expression in the gonads.
Results
Analysis of double-mutant mice
In the gonads of Kit-deficient W mutant mice, the degeneration of testicular germ cells and oocytes was reported to be accompanied by cell death.10 We investigated whether Fas is involved in these events because we and other groups previously showed the expression of Fas in both male and female gonads.31,32,33,34,35,36,37,38,41,43,44,45,46 To clarify whether Fas is involved in germ cell degeneration in the gonads of W mutant mice, we generated double-mutant mice (Wv/Wv:Fas−/−) by intercrossing the spontaneous Kit mutant Wv mice with Fas-deficient mice against the same C57BL/6 background.
We first examined whether the number of germ cells in the gonads of the mutant mice changes with the Fas gene status. The number of Wv/Wv homozygous newborns was less than that expected as a result of Mendel's laws, while a significant effect of Fas gene status was not observed. In addition, Wv/Wv homozygous mice often died within 2 weeks because of anemia (data not shown), while some mutant mice survived for more than 8 weeks. Their testes and ovaries could then be examined. We measured the size of the testes of the surviving Wv mutant mice, and found the testes of Wv/Wv homozygous mice to be smaller (approximately 10–20% in size) than those of Wv/+ or wild-type mice regardless of Fas status (data not shown).
Histological analysis of testes in the double-mutant mice
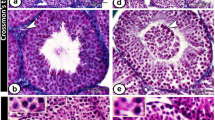
Next, we histologically examined the testes of the 8-week-old mutant mice by staining with hematoxylin and eosin (Figure 1). Testes of Wv/+:Fas−/− mice were histologically normal. Germ cells at all developmental stages including spermatogonia, primary spermatocytes, secondary spermatocytes, haploid spermatids and sperm were observed (Figure 1a,d). Fas-deficiency did not induce hyperplasia of germ cells in these mutant male mice. In the testes of Wv/Wv:Fas+/+ mice, seminiferous tubules were abnormal, resulting in no differentiated germ cells (Figure 1b,e), as described previously in Wv mutant mice.10 Interestingly, in the testes of Wv/Wv:Fas−/− mice, spermatocytes, spermatids and sperm were detected in several seminiferous tubules (Figure 1c,f), indicating that Fas deficiency can rescue testicular germ cells in Wv/Wv mice. As summarized in Table 1, about 90% of seminiferous tubules in Wv/Wv:Fas−/− mice had only Sertoli cells with or without a very few spermatogonia, while we observed a significant number of spermatocytes that differentiated into the pachytene stage in the other 10% of tubules. Further, differentiated spermatids and sperm could be also observed in about 5% of seminiferous tubules. The seminiferous tubules containing differentiated germ cells at the meiotic or a more advanced stage of spermatogenesis seemed to be located around rather than in the center of the testes of Wv/Wv:Fas−/− mice (Figure 1c). However, we could not detect sperm in the epididymis of Wv/Wv:Fas−/− mice by histological analysis (data not shown). Taking all the results into consideration, Fas was shown to be involved in the degeneration of testicular germ cells in Wv mutant mice lacking signals from Kit, while other signals might also play a role in the degeneration.

Histological analyses of testes from 8-week-old mutant mice. Sections prepared from testes of Wv/+:Fas−/− (a,d), Wv/Wv:Fas+/+ (b,e) and Wv/Wv:Fas−/− (c,f) mice were stained with hematoxylin and eosin. Photographs were taken under a phase-contrast microscope at × 32 (a–c) and × 160 (d–f). Arrows in (c) show the tubules containing differentiated germ cells indicated by arrowheads in (f). Scale bars correspond to 125 μm
We also examined the propagative activity of Wv/Wv:Fas−/− male mice. However, we observed no fertility in Wv/Wv:Fas−/− male mice mating with wild-type females. The number of undegenerated germ cells and/or differentiated sperm in the double-mutant mice might be too small for fertilization.
Histological analysis of ovaries in the double-mutant mice
Female gonads were also analyzed in Wv/+:Fas−/−, Wv/Wv:Fas+/+ and Wv/Wv:Fas−/− mice. Figure 2 shows the results of a histological analysis of ovaries in 4-week-old animals. Ovaries of Wv/+:Fas−/− mice displayed a normal histology, where oocytes and follicles at various stages of development were detected (Figure 2a,d). This phenotype of Wv/+:Fas−/− mice is similar to that of lpr mice, which exhibit a spontaneous loss of function in the Fas gene. On the other hand, ovaries of Wv/Wv:Fas+/+ mice were shown to be only very small (Figure 2b,e), with no oocytes or follicles. Importantly, in ovaries of Wv/Wv:Fas−/− mice, we detected primary follicles, growing follicles and Graffian follicles with oocytes (Figure 2c,f). Thus, Kit deficiency affected the gametogenesis of female gonads more than male gonads in younger mice, and inactivation of Fas resulted in the diminution of the degeneration of oocytes and follicles in the Kit-deficient ovaries. Results for ovaries are summarized in Table 2. They clearly indicate that loss of function of Fas prevents the degeneration of germ cells in both male and female gonads of Wv/Wv mice.

Histological analyses of ovaries isolated from 4-week-old mutant mice. Sections of ovaries of Wv/+:Fas−/− (a,d), Wv/Wv:Fas+/+ (b,e) and Wv/Wv:Fas−/− (c and f) mice were stained with hematoxylin and eosin. Photographs were taken under a phase-contrast microscope at × 32 (a–c) and × 80 (d–f). Scale bars correspond to 125 μm
Although Wv/Wv:Fas−/− female mice were examined in terms of their fertility, they did not become pregnant (data not shown). In addition, oocytes of Wv/Wv:Fas−/− mice were shown to be impaired with aging, and most of the oocytes degenerated in the ovary of 8-week-old Wv/Wv:Fas−/− female mice (data not shown). All the results indicate that Fas is involved in the degeneration of oocytes in 4-week-old Wv/Wv:Fas−/− female mice, while mechanisms other than Fas-mediated apoptosis might play a role in the infertility and the degeneration of oocytes of 8-week-old Wv/Wv:Fas−/− mice.
In vivo blocking of Kit signals by anti-Kit mAb causes germ cell degeneration in wild-type but not Fas-deficient mice
We next examined the effects of Fas deficiency on the degeneration of testicular germ cells in another mouse system with suppressed Kit signals. We artificially blocked the Kit signaling pathway through injection of an antagonistic anti-c-kit mAb ACK2 into wild-type C57BL/6 and Fas-deficient mice. We analyzed the population of testicular germ cells from both mice after injection of ACK2. It has been demonstrated that differentiating spermatogonia and early preleptotene spermatocytes expressing Kit degenerated after injection of ACK2 which can block the function of Kit.18 We examined the testes of the antibody-treated animals by an immunohistochemical method with an anti-germ cell-specific mAb, TRA98, and stained another serial section with Hoechst (Figure 3). At 48 h after injection of ACK2, most differentiating type A and intermediate spermatogonia had disappeared in the wild-type testes (Figure 3a), although differentiated germ cells and interstitial cells were observed in Hoechst-stained sections (Figure 3c). In contrast, many spermatogonia still existed even after 48 h in Fas-deficient mice (Figure 3b). These results supported our results for Wv/Wv:Fas−/− double-mutant male mice (Figure 1 and Table 1) and strongly indicate that Fas is involved in testicular germ cell degeneration induced by defects of Kit signaling.

Immunohistochemistry of testes from ACK2-injected adult mice. Frozen sections were prepared from testes of wild-type C57BL/6 mice (a,c) and Fas−/− mice (b,d), to which anti-c-kit mAb ACK2 was administered 48 h earlier, as described in Materials and Methods. The section was then stained with the germ cell-specific mAb TRA98 (a,b), and another serial section was stained with Hoechst (c,d). Experiments were repeated five times using different mice and the data presented show representative results. Photographs were taken under a phase-contrast microscope at × 160. Arrows and arrowheads show spermatogonia and differentiated haploid spermatids, respectively. Scale bars correspond to 25 μm
Expression of Fas in testes from wild-type and homozygous Wv/Wv mutant mice
Our data thus suggest that Kit-mediated signals inhibit the Fas-mediated apoptosis-inducing signal. To clarify the molecular mechanism of the Kit-induced inhibition, we analyzed the expression level of Fas because activation of Ras, which must occur downstream of Kit, was previously reported to decrease the expression.48 In our case, we speculated that the deficiency of Kit signaling might cause the induction of Fas expression on the germ cells. At first, we analyzed the expression at the RNA level by Northern blot hybridization. We examined total RNA isolated from the testes of 6-week-old wild-type C57BL/6 and homozygous Wv/Wv mutant mice (Figure 4). Fas transcripts were significantly detected in the testes from Wv/Wv mutant mice, but we could not detect Fas mRNA in the testes of wild-type mice. This result suggested that Fas is expressed on spermatogonia and/or interstitial cells. Alternatively, the difference in Fas expression might reflect the difference of the cell population in the testes of wild-type and Wv/Wv mutant mice.

Northern blot analysis of Fas transcripts in testes. Northern blotting was carried out with total RNA isolated from testes of 5- to 6-week-old wild-type C57BL/6 and Wv/Wv mutant mice as described in Materials and Methods using 32P-labeled mouse Fas (upper panel) and human EF1-α (lower panel) cDNAs as DNA probes.
To identify the cell surface expression of Fas, we performed flow cytometric analysis with the anti-Fas mAb Jo-2. Fas expression was examined on the surface of testicular cells prepared from 3- to 4-week-old wild-type C57BL/6 and homozygous Wv/Wv mutant mice. In wild-type testes, few cells expressing Fas were observed, while such cells were detected in Wv/Wv mutant mice (Figure 5a). Then, we carried out a two-dimensional flow cytometric analysis with the Jo-2 and EE2 mAbs in order to clarify whether Fas is expressed on spermatogonia or not. Previous study demonstrated the expression of EE2 antigen on the germ cell surface; the expression was predominant prior to meiosis, and gradually decreased as the differentiation proceeded.49 EE2-bright cells correspond to type A and primitive spermatogonia, while EE2-intermediate cells correspond to intermediate and type B spermatogonia, preleptotene, leptotene and zygotene spermatocytes. EE2-negative cells correspond to pachytene spermatocytes or cells in the more advanced stages of spermatogenesis and interstitial cells. Flow cytometric analysis clearly indicated that Fas was significantly expressed on EE2-positive cells in Wv/Wv mutant mice although little expression of Fas was detected on EE2-positive cells in wild-type mice (Figure 5b). We then examined germ cells of 2-week-old mice, and found the induction level of Fas expression in 2-week-old than 3- to 4-week-old Wv/Wv mutant mice (Figure 5b). Fas expression was indicated to be upregulated in testicular germ cells of 3- to 4-week-old mice mainly because of a deficiency of Kit.

Flow cytometric analyses of Fas expression on mouse testicular germ cells. (A) Single-cell suspensions isolated from testes of 3- to 4-week-old wild-type C57BL/6 mice (left panel) and Wv/Wv mutant mice (right panel) were stained with (solid lines) or without (dotted lines) FITC anti-Fas mAb Jo-2. (B) Testicular cells of 3- to 4-week-old or 2-week-old wild-type C57BL/6 mice and Wv/Wv mutant mice were stained with PE-anti-testicular germ cell mAb EE2, together with (right panels) or without (left panels) FITC-Jo-2. (C) Single-cell suspensions isolated from testes of 3- to 4-week-old wild-type C57BL/6 mice (left panel), Wv/Wv mutant mice (center panel) and Wv/Wv:Fas−/− mice (right panel) were stained with EE2 and ACK2. The squares, indicated as a, d and g, correspond to spermatogonia; b, e and h to primitive spermatogonia; and c, f and i to interstitial cells and well-differentiated germ cells. (D) Fas expression on testicular cells in the squares in (C). Cells were stained with (solid lines) or without (dotted lines) FITC Jo-2. All experiments were repeated five times, and the data presented show representative results
To clearly distinguish expression levels of Fas between SCF-dependent and -independent spermatogonia, we carried out flow cytometric analyses by triple staining with EE2, ACK2 and Jo-2. Figure 5c shows the results of two-dimensional flow cytometric analyses with EE2 and ACK2 in wild-type (left panel), Wv/Wv mutant (center panel) and Wv/Wv:Fas−/− double-mutant (right panel) mice. Kit-positive and EE2-positive cells (squares a, d and g) correspond to spermatogonia that require SCF-induced Kit signals for differentiation. Kit-negative and EE2-bright cells (squares b, e and h) correspond to primitive spermatogonia such as testicular stem cells which have the ability to self-proliferate. Kit-negative and EE2-negative cells (squares c, f and i) correspond to Sertoli cells and pachytene spermatocytes or more advanced stages of spermatogenesis. In Wv/Wv mutant mice, primitive spermatogonia, encircled with the square e, were decreased in number when compared with those of wild-type mice. In Wv/Wv:Fas−/− double-mutant mice, however, a drastic recovery of primitive spermatogonia was not observed. Although the percentage of wild-type spermatogonia requiring Kit signals for differentiation, encircled with the square a (containing 10% of total cells), was about one-third of that of Wv/Wv spermatogonia (square d, 24%), the total cell number of spermatogonia/testis in wild-type mice was more than six times that in Wv/Wv mutant mice, because total cell number/testis for cytofluorometric analysis in wild-type mice was more than 20 times that in Wv/Wv mutant mice. Interestingly, Fas deficiency increased the number of spermatogonia requiring Kit signals for differentiation more than two-fold when compared with that of Wv/Wv mutant mice, because (1) the percentage of spermatogonia requiring Kit signals for differentiation in Wv/Wv and Wv/Wv:Fas−/− mutant mice was comparable (square d, 24%, and square g, 29%, respectively) and (2) total cell number/testis for cytofluorometric analysis in Wv/Wv:Fas−/− mice was more than two times that in Wv/Wv mutant mice.
Next, we examined Fas expression on Kit-positive and EE2-positive cells, Kit-negative and EE2-bright cells and Kit-negative and EE2-negative cells encircled with the squares a–f in Figure 5c (Figure 5d). Wv/Wv mutant mice were shown to significantly express Fas in all the testicular cells, while wild-type mice expressed little Fas in any testicular cells.
All the results indicated that the expression of Fas on germ cells was upregulated in the testes of Wv/Wv mutant mice, and germ cell degeneration induced by defects in the Kit signaling system was suggested to be caused by activation of the Fas-mediated signaling pathway.
SCF downregulates Fas expression on Kit-expressing NIH3T3 cells
To confirm the downregulation of Fas by Kit-induced signals, we examined the effects of SCF on Kit-expressing cells in vitro. We transfected c-kit cDNA into NIH3T3 cells and established several Kit-positive stable clones. Then, we analyzed the expression of Fas on these clones by flow cytometry after treatment with 10 μg/ml of mouse recombinant SCF for 48 h. A significant decrease of Fas was detected on the Kit-expressing stable clone NIH-K11 but not parental NIH3T3 cells after treatment with SCF (Figure 6a,b). The other Kit-positive stable clones showed essentially similar characteristics to those of NIH-K11 cells (data not shown). These results directly show that the Kit signal induces the downregulation of Fas.

Flow cytometric analyses of Fas expression on SCF-treated fibroblasts expressing exogenous Kit. Parental NIH3T3 (a) and Kit-expressing stable clone NIH-K11 (b) cells were stimulated with (bold lines) or without (fine lines) 10 ng/ml of SCF for 48 h and then stained with (dotted lines) or without (solid lines) FITC-Jo-2. NIH-K11 cells were also stimulated with SCF for 48 h in the presence of 100 nM wortmannin (c) or 10 μM U0126 (d). Cells were pretreated with each inhibitor for 30 min prior to stimulation with SCF. Experiments were repeated five times, and the data presented show representative results
We then examined which downstream signal of Kit plays a role in the downregulation of Fas expression. Since the downregulation was speculated to be induced by survival signals mediated through Kit, we focused on two signaling pathways downstream of Kit, PI3K/Akt and Ras/MEK/ERK. Activation of the Ras/MEK/ERK pathway was reported to inhibit Fas-mediated apoptosis,50 while activation of PI3K/Akt pathway was suggested to suppress the expression level of cell surface Fas.48 In addition, Kit-mediated activation of the PI3K/Akt pathway was demonstrated to be significant in gametogenesis.51,52 We analyzed the effects of two kinds of inhibitors that specifically prevent Kit-mediated signaling: the PI3K-specific inhibitor wortmannin and the MEK1/2-specific inhibitor U0126. These inhibitors were added to the culture medium of NIH-K11 cells together with SCF and Fas expression was examined 48 h later with a flow cytometer (Figure 6c and d). Downregulation of Fas was suppressed by the treatment with wortmannin, while U0126 showed no significant effect. Similar results were obtained with the PI3K inhibitor LY294002 and MEK1 inhibitor PD98059 (data not shown). Thus, PI3K inhibitors but not MEK inhibitors blocked the downregulation of Fas by SCF.
To biochemically detect the activation of downstream signaling molecules of Kit, which was inhibited by wortmannin or U0126, we examined the phosphorylation of Akt and ERK, which are downstream molecules of PI3K and MEK, respectively. Immunoblot analyses for Akt and ERK were carried out using antiphospho-Akt (Thr308) and antiphospho-p44/42 MAPK antibodies, respectively (Figure 7). After 15 min stimulation of NIH-K11 (lanes 1–4) and NIH3T3 cells (lanes 5, 6) with 10 ng/ml of SCF, the phosphorylation of Akt and ERK was observed in SCF-stimulated NIHK-11 cells (lane 2), but in neither unstimulated NIH-K11 (lane 1) nor parental NIH3T3 cells (lanes 5, 6). In addition, the SCF-induced phosphorylation of Akt and ERK was canceled by addition of wortmannin (lane 3) and U0126 (lane 4), respectively. Thus, the SCF/Kit-induced downregulation of Fas was shown to correlate well with the activation of PI3K/Akt.

Immunoblot analyses of SCF-treated NIHK-11 and NIH3T3 cells. Cells were precultured without serum for 12 hr before treatment with 10 ng/ml of SCF together with or without wortmannin or U0126. Cells were pretreated with each inhibitor for 30 min prior to SCF stimulation. Akt, Akt phosphorylated at Thr 308 and phosphorylated MAPK (ERK) were detected with anti-Akt (upper panel), antiphospho Akt (center panel) and anti-phospho MAPK (lower panel) antibodies, respectively
Akt plays an important role in downregulation of Fas expression on NIH3T3 cells
Figures 6 and 7 clearly show that PI3K is involved in the regulation of Fas expression. To clarify the downstream signaling molecule of PI3K regulating Fas expression, we transfected a constitutively active Akt (caAkt; HA-mΔ4–129Akt) into NIH3T3 cells and analyzed Fas expression by flow cytometry (Figure 8). GFP-expression vector was cotransfected with a caAkt or control vector, and Fas expression in GFP-positive cells was analyzed. The expression level of Fas on caAkt transfectants was significantly lower than that of cells transfected with the control vector. Thus, activation of Akt downregulates Fas expression on NIH3T3 cells.

Effect of constitutive active Akt on expression of Fas. Control vector (upper panels) or the expression vector for constitutive active Akt (lower panels) was cotransfected with the expression vector for GFP into NIH3T3 cells. After 48 hr cultivation, cells were stained with control-PE-IgG (left panels) or PE-conjugated Jo-2 (right panels), and analyzed by flow cytometry. The proportion (%) of separated respective cells into four parts is shown in the figure. Experiments were repeated five times, and the data presented show representative results
Taking these results together, we conclude that stimulation of the Kit receptor downregulates Fas expression by activating the PI3K/Akt pathway in not only exogenous Kit-expressing fibroblasts in vitro but also germ cells in gonads in vivo.
Discussion
In this study, we demonstrated that a failure of Kit signaling induces activation of the Fas system in both male and female gonads by analyzing Wv/Wv, Fas−/− and double-mutant mice. Further, we verified that the PI3K/Akt signaling pathway downstream of Kit regulates expression of Fas. From this evidence, we proposed that crosstalk occurs between Kit-mediated and Fas-mediated signaling pathways in the mouse gonads.
In the male gonads of Wv/Wv:Fas−/− mice, we observed differentiating germ cells in approximately 10% of seminiferous tubules, but could not detect a recovery of germ cells in all seminiferous tubules (Figure 1). These results indicate that Fas is necessary but not sufficient to induce germ cell apoptosis in Wv mutant mice, and other factors must be involved in the loss of germ cells in Wv mutant mice. Recently, a deficiency of p53 was reported to rescue germ cell degeneration in Wv/Wv mice.53 Although these Wv/Wv:p53−/−double-mutant mice showed propagative activity, their sperm counts were less than 7% of those in wild-type mice. Inactivation of p53 may not be sufficient to rescue germ cell degeneration induced by a failure of Kit signaling. We suppose that both Fas and p53 work cooperatively as an executioner in germ cell death. Our hypothesis is supported by a previous report describing that Fas was responsible for the p53-independent germ cell apoptosis in response to heat stress in lpr/lpr:p53−/− double-mutant mice.54 Thus, both Fas- and p53-dependent signaling pathways play a role in the apoptosis of germ cells.
Interestingly, a deficiency of p53 did not rescue oocytes of Wv/Wv mice from degeneration.53 In contrast, our Wv/Wv:Fas−/− mice displayed an apparently normal oogenesis and folliculogenesis even though the Kit signaling was defective (Figure 2). These results indicate that loss of function of Fas, but not that of p53, leads to the recovery of both oocytes and granulosa cells in Wv mutant mice. Many previous reports, including ours, demonstrated expression of Fas on granulosa cells.31,32,33,34,35,36,37,38 A few reports suggested expression of Fas on oocytes32,38,39 although we could not detect it. On the other hand, Kit expression was shown to be restricted on oocytes but not on granulosa cells in follicles,13,18,55 while deficiency of Kit leads to Fas-induced loss of granulosa cells as well as oocytes. We wondered why Kit-positive and Fas-negative oocytes and Kit-negative and Fas-positive granulosa cells are similarly degenerated in Wv mutant mice. In oocytes, failure of Kit signaling might induce Fas expression. To explain the loss of granulose cells, two previous reports are helpful, which insisted on the presence of paracrine factor(s) moving between oocytes and granulosa cells in the follicles.55,56 Loss of the paracrine factor(s) from oocytes in Kit-defective mice may activate Fas-mediated apoptotic signals in granulosa cells.
Kit signals are essential not only in gametogenesis but also in hematopoiesis and melanogenesis. Therefore, we examined bone marrow cells isolated from Wv/Wv:Fas+/+ and Wv/Wv:Fas−/− mice, but we could not observe a noticeable difference in their cell populations by flow cytometric analyses (data not shown). Moreover, Fas expression was previously reported on granulocytes expressing Kit.57 Thus, Fas expression is not regulated by Kit-mediated signals in hematopoietic cells. In addition, our double-mutant mice displayed the same coat color as Wv/Wv:Fas+/+ mice, suggesting that inactivation of Fas has no significant effect on the survival of melanocytes. Thus, the downregulation of Fas expression by Kit-mediated signals is suggested to be a cell type-specific event. This might explain the moderate level of SCF-induced downregulation of Fas in Kit-expressing fibroblasts (Figure 6).
In this study, we demonstrated the existence of crosstalk between Fas-mediated apoptotic signals and Kit-mediated survival signals. In vivo, Fas expression was upregulated on Kit-deficient Wv spermatogonia (Figure 5). On 4-week-old Wv spermatogonia, which soon undergo apoptosis, the expression of Fas is strong, while on 2-week-old Wv spermatogonia, which survive for some time, it is negligible. These results correlate well with the germ cell apoptosis associated with the aging of Wv/Wv mutant mice. From the results of two-dimensional flow cytometric analyses with EE2 and ACK2, the number of primitive spermatogonia was shown to dramatically decrease in Wv/Wv mutant mice. While primitive spermatogonia of Wv/Wv mutant mice expressed Fas, the number of these cells in Wv/Wv:Fas−/− double-mutant mice recovered only slightly. It is considered that Fas may not be directly involved in the degeneration of primitive spermatogonia in Wv/Wv mutant mice, and other factor(s) might contribute to the loss. In contrast, the number of SCF-requiring spermatogonia recovered in Wv/Wv:Fas−/− double-mutant mice compared with Wv/Wv mutant mice. All the results indicated that a defect in Fas can rescue SCF-dependent spermatogonia from apoptosis, resulting in an increase in the number of more differentiated cells, although Fas deficiency cannot rescue primitive spermatogonia.
In addition, we also showed that Fas is downregulated in vitro on stimulation with SCF in Kit-expressing NIH3T3 cells (Figure 6). Our in vitro analyses with inhibitors for PI3K or transfectants expressing caAkt clearly verified the correlation between the activation of Akt and downregulation of Fas expression (Figures 6,7 and 8). The significance of the Kit-mediated PI3K/Akt signaling pathway in gametogenesis was previously demonstrated by analyzing mutant Kit knockin mice.51,52 These mutant mice, expressing mutated Kit at Tyr719 which cannot interact with the p85 subunit of PI3K, were shown to be defective in spermatogenesis and oogenesis but not in hematopoiesis and melanogenesis. These results coincided well with our results in this study. In addition, Akt1-deficient mice were recently generated and showed a similar phenotype to that observed in the knockin mice of the Kit mutant.58 Thus, the PI3K/Akt signaling pathway via Kit is essential for the survival of testicular germ cells and oocytes, which suppresses Fas-mediated apoptotic signaling by downregulating the expression of Fas in the gonads.
In this study, we demonstrated that Fas expression is downregulated via the activation of PI3K/Akt signaling. This result coincided with a previous report indicating downregulation of Fas on the activation of Ras/PI3K/Akt in NIH3T3 cells.48 However, several lines of evidence indicated that the PI3K/Akt signaling pathway negatively controls apoptosis by regulating multiple signaling molecules. By inactivating a forkhead transcription factor, FKHRL-1, Akt was reported to downregulate Fas ligand and upregulate expression of c-FLIP, which is known as an antiapoptotic molecule against Fas-induced apoptosis.59,60 As we have not shown the regulation of these target molecules by activated PI3K/Akt in this study, it is necessary to clarify them with further studies.
We used NIH3T3 cells to investigate the downregulation of Fas by Kit-mediated and caAkt-induced signals (Figures 6 and 8). Further, Ras and its downstream molecule PI3K were reported to downregulate Fas expression, although activation of Akt was not shown.48 However, we could not detect a noticeable reduction of endogenous Fas expression on Balb3T3 cells expressing caAkt as described in a previous report, where we showed that Fas-mediated apoptosis was strongly inhibited by treatment with basic FGF, a ligand for another RTK, through inhibiting intracellular signals via Fas by activation of ERK.50 These results indicate that inhibitory mechanisms for Fas-mediated apoptosis are regulated by multiple signaling molecules including Akt and ERK, which affect not only the Fas expression level but also the intracellular signaling pathway via Fas. Kit-mediated signals might inhibit Fas-mediated apoptosis both in vivo and in vitro by not only suppressing Fas expression but also blocking the intracellular signaling pathway via Fas.
In this study, we revealed the negative regulation of Fas-mediated signaling by survival signals induced by Kit/SCF through in vivo and in vitro analyses of Wv mutant mice and fibroblasts, respectively. Our results provide a clue to the relation between various survival signals and death signals.
Materials and Methods
Materials and antibodies
The nylon membrane Hybond-N was purchased from Amersham Bioscience, PI3K inhibitors wortmannin and LY294002 from SIGMA and Cell Signaling, respectively, the MEK1/2 inhibitor U0126 and MEK1 inhibitor PD98059 from Cell Signaling and the polyvinylidene difluoride (PVDF) membrane Immobilon from Millipore.
Monoclonal antibodies EE249 and TRA9861 that recognize testicular germ cells and anti-c-kit mAb ACK262 were prepared as described previously. Fluorescein isothiocyanate (FITC)- and phycoerythrin (PE)-conjugated anti-Fas mAb Jo-2 were purchased from BD-Pharmingen. Horseradish peroxidase (HRP)-conjugated anti-Rat IgG Ab was from Cappel, biotin-conjugated anti-rat κ+λ light chains mAb from Sigma, HRP-conjugated anti-mouse Ig from Amersham Bioscience, PE- and APC-conjugated avidin from BD-Pharmingen and PE-conjugated ACK2 from eBioscience. Anti-Akt mAb, anti-Phospho-Akt (Thr308) Ab and anti-Phospho-p44/42 MAPK mAb and HRP-linked anti-rabbit IgG Ab were purchased from Cell Signaling.
Mice
Wv/Wv:Fas−/− mice were generated by intercrossing heterozygous Wv/+ mutant mice (SLC, Shizuoka, Japan) with Fas-deficient mice63 having the same C57B/6 background. A Fas-deficient C57B/6 background was generated by backcrossing seven times with C57B/6 mice. The genotype of the Fas mutants was identified by PCR assay with DNA isolated from tails. The Wv mutation was distinguished by coat color. For the dissection of reproductive organs, mice were killed using carbon dioxide gas.
Histological analysis
Testes and ovaries dissected from mice were fixed overnight in Bouin's solution, embedded in paraffin after dehydration, and cut at 5 μm. Sections were stained with hematoxylin and eosin according to standard procedures.
Administration of anti-c-kit mAb ACK2
To block c-kit function, 100 μg/mouse of purified anti-c-kit mAb ACK2 was intravenously injected into adult (8- to 12-week-old) mice as described previously.18 For a control, the same volume of phosphate-buffered saline (PBS) was injected.
Immunohistochemistry
Dissected testes were embedded in an O.C.T compound (Tissue-Tec, Sakura, Tokyo, Japan), frozen quickly in liquid nitrogen, and sectioned at 8 μm with a cryostat (Carl Zeiss). Sections were fixed for 30 min in PBS containing 4% paraformaldehyde at room temperature, and then stained with the germ cell-specific mAb TRA98 as follows. The sections were preincubated in PBS containing 1% goat serum for 1 h, and then incubated with TRA98 in PBS containing 3% skimmed milk and 0.1% Tween 20 (PBS-MT) at room temperature for 1 h. After three washes with PBS containing 0.1% Tween 20 (PBS-T), the sections were incubated with HRP-conjugated anti-Rat IgG (1:1000) in PBS-MT for 2 h. After three more washes with PBS-T, samples were developed using a metal-enhanced diaminobendizine (DAB) substrate kit (PIERCE).
Northern blot analysis
Cellular total RNA was prepared from testes of C57BL/6 and Wv/Wv mutant mice using ISOGEN (Nippon Gene, Tokyo, Japan) according to the manufacturer's instructions. Total RNA (20 μg) was loaded on formaldehyde–agarose gel for electrophoresis and transferred to nylon membrane. DNA probes for mouse Fas and EF1-α were labeled with [α-32P] dCTP (specific activity >3000 Ci/mmol, ICN) using an RTG-DNA labeling kit (Amersham Bioscience) with full-length mouse Fas cDNA26 and full-length EF1-α cDNA,64 respectively, and purified using a NICK Column (Pharmacia Biotech) according to the manufacturer's instructions. The membrane was hybridized with probes using PerfectHyb Hybridization Solution (TOYOBO, Osaka, Japan) at 68°C overnight, and washed with 2 × SSC/0.1% SDS for 5 min twice at 68°C and then 0.1 × SSC/0.1% SDS for 30 min twice at 68°C. The membrane was developed by placing it in contact with X-ray film.
Flow cytometric analysis
Testes dissected from 2- to 6-week-old wild-type C57B/6 and Wv/Wv:Fas+/+ mice were cut into small pieces with a razor blade and pipetted several times in PBS to isolate testicular cells, and the testicular single-cell suspension was collected through a nylon mesh. Aliquots of 1 × 106 cells were incubated for 1 h at 4°C with or without 1 μg of FITC-conjugated anti-Fas mAb Jo-2 in flow cytometric buffer (PBS containing 5% FCS and 0.04% sodium azide). In the case of double staining with both EE2 and Jo-2 or triple staining with EE2, ACK2 and Jo-2, cells were initially incubated with an EE2 mAb for 1 h at 4°C, then with a biotin-conjugate anti-rat κ+λ light chain mAb for 30 min. After blocking with rat IgG for 30 min, cells were finally incubated with PE-conjugated avidin and FITC-conjugated Jo-2 or APC-conjugated avidin, PE-conjugated ACK2 and FITC-conjugated Jo-2 for 1 h. NIH3T3 and its derivative NIH-K11 were stimulated with mouse recombinant SCF (Calbiochem) with or without wortmannin, LY294002, U0126 or PD98059 for 48 h. Cells were then scraped with a rubber policeman and stained with Jo-2 as described above. All the samples, rinsed with flow cytometric buffer twice and fixed with PBS containing 3.5% formaldehyde, were analyzed by flow cytometry on an EPICS Elite (Beckman Coulter).
Establishment of stable transfectants
Mouse c-kit cDNA subcloned into the vector pcDNA3 (Invitrogen) was provided by Drs. Ueda and Kanakura (Osaka University) and was introduced into NIH3T3 cells using Lipofectamin Plus reagent (Invitrogen). After selection with G418 and subsequent cloning, clones stably expressing Kit were established.
Western blot analysis
Total cell lysates of NIH3T3 cells and transfectants were separated by SDS-PAGE and transferred to PVDF membranes. Then, the membranes were immersed in Tris-buffered saline (pH 7.6) with 0.1% Tween 20 (TBS-T), 5% skimmed milk and 2% FCS at room temperature for 1 h and incubated with the primary antibodies at room temperature for 1 h. The membranes were washed with TBS-T three times, and incubated with the secondary antibodies at room temperature for 1 h. Specific signals were detected using the enhanced Western Lightning (PerkinElmer Life Science) detection system on X-ray film.
Abbreviations
- ERK:
-
extracellular signal-regulated kinase
- MEK:
-
MAP kinase-ERK kinase
- PI3K:
-
phosphatidylinositol 3-phosphate kinase
- RTK:
-
receptor tyrosine kinase
- SCF:
-
stem cell factor
References
Qiu FH, Ray P, Brown K, Barker PE, Jhanwar S, Ruddle FH and Besmer P (1988) Primary structure of c-kit: relationship with the CSF-1/PDGF receptor kinase family–oncogenic activation of v-kit involves deletion of extracellular domain and C terminus. EMBO J. 7: 1003–1011
Besmer P, Murphy JE, George PC, Qiu FH, Bergold PJ, Lederman L, Snyder Jr HW, Brodeur D, Zuckerman EE and Hardy WD (1986) A new acute transforming feline retrovirus and relationship of its oncogene v-kit with the protein kinase gene family. Nature 320: 415–421
Yarden Y, Kuang WJ, Yang-Feng T, Coussens L, Munemitsu S, Dull TJ, Chen E, Schlessinger J, Francke U and Ullrich A (1987) Human proto-oncogene c-kit: a new cell surface receptor tyrosine kinase for an unidentified ligand. EMBO J. 6: 3341–3351
Chabot B, Stephenson DA, Chapman VM, Besmer P and Bernstein A (1988) The proto-oncogene c-kit encoding a transmembrane tyrosine kinase receptor maps to the mouse W locus. Nature 335: 88–89
Geissler EN, Ryan MA and Housman DE (1988) The dominant-white spotting (W) locus of the mouse encodes the c-kit proto-oncogene. Cell 55: 185–192
Copeland NG, Gilbert DJ, Cho BC, Donovan PJ, Jenkins NA, Cosman D, Anderson D, Lyman SD and Williams DE (1990) Mast cell growth factor maps near the steel locus on mouse chromosome 10 and is deleted in a number of steel alleles. Cell 63: 175–183
Huang E, Nocka K, Beier DR, Chu TY, Buck J, Lahm HW, Wellner D, Leder P and Besmer P (1990) The hematopoietic growth factor KL is encoded by the Sl locus and is the ligand of the c-kit receptor, the gene product of the W locus. Cell 63: 225–233
Zsebo KM, Williams DA, Geissler EN, Broudy VC, Martin FH, Atkins HL, Hsu RY, Birkett NC, Okino KH and Murdock DC (1990) Stem cell factor is encoded at the Sl locus of the mouse and is the ligand for the c-kit tyrosine kinase receptor. Cell 63: 213–224
Russell ES (1979) Hereditary anemias of the mouse: a review for geneticists. Adv. Genet. 20: 357–459
Besmer P, Manova K, Duttlinger R, Huang EJ, Packer A, Gyssler C and Bachvarova RF (1993) The kit-ligand (steel factor) and its receptor c-kit/W: pleiotropic roles in gametogenesis and melanogenesis. Development, Suppl.: 125–137
Nocka K, Tan JC, Chiu E, Chu TY, Ray P, Traktman P and Besmer P (1990) Molecular bases of dominant negative and loss of function mutations at the murine c-kit/white spotting locus: W37, Wv, W41 and W. EMBO J. 9: 1805–1813
Reith AD, Rottapel R, Giddens E, Brady C, Forrester L and Bernstein A (1990) W mutant mice with mild or severe developmental defects contain distinct point mutations in the kinase domain of the c-kit receptor. Genes Dev. 4: 390–400
Manova K, Nocka K, Besmer P and Bachvarova RF (1990) Gonadal expression of c-kit encoded at the W locus of the mouse. Development 110: 1057–1069
Manova K, Huang EJ, Angeles M, De Leon V, Sanchez S, Pronovost SM, Besmer P and Bachvarova RF (1993) The expression pattern of the c-kit ligand in gonads of mice supports a role for the c-kit receptor in oocyte growth and in proliferation of spermatogonia. Dev. Biol. 157: 85–99
Rossi P, Dolci S, Albanesi C, Grimaldi P, Ricca R and Geremia R (1993) Follicle-stimulating hormone induction of steel factor (SLF) mRNA in mouse Sertoli cells and stimulation of DNA synthesis in spermatogonia by soluble SLF. Dev. Biol. 155: 68–74
Sandlow JI, Feng HL and Sandra A (1997) Localization and expression of the c-kit receptor protein in human and rodent testis and sperm. Urology 49: 494–500
Sorrentino V, Giorgi M, Geremia R, Besmer P and Rossi P (1991) Expression of the c-kit proto-oncogene in the murine male germ cells. Oncogene 6: 149–151
Yoshinaga K, Nishikawa S, Ogawa M, Hayashi S, Kunisada T and Fujimoto T (1991) Role of c-kit in mouse spermatogenesis: identification of spermatogonia as a specific site of c-kit expression and function. Development 113: 689–699
Schlessinger J (2000) Cell signaling by receptor tyrosine kinases. Cell 103: 211–225
Linnekin D (1999) Early signaling pathways activated by c-Kit in hematopoietic cells. Int. J. Biochem. Cell B 31: 1053–1074
Ashkenazi A and Dixit VM (1999) Apoptosis control by death and decoy receptors. Curr. Opin. Cell Biol. 11: 255–260
Yonehara S, Ishii A and Yonehara M (1989) A cell-killing monoclonal antibody (anti-Fas) to a cell surface antigen co-downregulated with the receptor of tumor necrosis factor. J. Exp. Med. 169: 1747–1756
Oehm A, Behrmann I, Falk W, Pawlita M, Maier G, Klas C, Li-Weber M, Richards S, Dhein J and Trauth BC (1992) Purification and molecular cloning of the APO-1 cell surface antigen, a member of the tumor necrosis factor/nerve growth factor receptor superfamily. Sequence identity with the Fas antigen. J. Biol. Chem. 267: 10709–10715
Suda T, Takahashi T, Golstein P and Nagata S (1993) Molecular cloning and expression of the Fas ligand, a novel member of the tumor necrosis factor family. Cell 75: 1169–1178
Rouvier E, Luciani MF and Golstein P (1993) Fas involvement in Ca(2+)-independent T cell-mediated cytotoxicity. J. Exp. Med. 177: 195–200
Watanabe-Fukunaga R, Brannan CI, Itoh N, Yonehara S, Copeland NG, Jenkins NA and Nagata S (1992) The cDNA structure, expression, and chromosomal assignment of the mouse Fas antigen. J. Immunol. 148: 1274–1279
Yonehara S, Nishimura Y, Kishil S, Yonehara M, Takazawa K, Tamatani T and Ishii A (1994) Involvement of apoptosis antigen Fas in clonal deletion of human thymocytes. Int. Immunol. 6: 1849–1856
Nagata S and Golstein P (1995) The Fas death factor. Science 267: 1449–1456
Nishimura Y, Ishii A, Kobayashi Y, Yamasaki Y and Yonehara S (1995) Expression and function of mouse Fas antigen on immature and mature T cells. J. Immunol. 154: 4395–4403
Suda T, Okazaki T, Naito Y, Yokota T, Arai N, Ozaki S, Nakao K and Nagata S (1995) Expression of the Fas ligand in cells of T cell lineage. J. Immunol. 154: 3806–3813
Leithauser F, Dhein J, Mechtersheimer G, Koretz K, Bruderlein S, Henne C, Schmidt A, Debatin KM, Krammer PH and Moller P (1993) Constitutive and induced expression of APO-1, a new member of the nerve growth factor/tumor necrosis factor receptor superfamily, in normal and neoplastic cells. Lab. Invest. 69: 415–429
Guo MW, Mori E, Xu JP and Mori T (1994) Identification of Fas antigen associated with apoptotic cell death in murine ovary. Biochem. Biophys. Res. Commun. 203: 1438–1446
Quirk SM, Cowan RG, Joshi SG and Henrikson KP (1995) Fas antigen-mediated apoptosis in human granulosa/luteal cells. Biol. Reprod. 52: 279–287
Hakuno N, Koji T, Yano T, Kobayashi N, Tsutsumi O, Taketani Y and Nakane PK (1996) Fas/APO-1/CD95 system as a mediator of granulosa cell apoptosis in ovarian follicle atresia. Endocrinology 137: 1938–1948
Sakamaki K, Yoshida H, Nishimura Y, Nishikawa S, Manabe N and Yonehara S (1997) Involvement of Fas antigen in ovarian follicular atresia and luteolysis. Mol. Reprod. Dev. 47: 11–18
Kim JM, Boone DL, Auyeung A and Tsang BK (1998) Granulosa cell apoptosis induced at the penultimate stage of follicular development is associated with increased levels of Fas and Fas ligand in the rat ovary. Biol. Reprod. 58: 1170–1176
Roughton SA, Lareu RR, Bittles AH and Dharmarajan AM (1999) Fas and Fas ligand messenger ribonucleic acid and protein expression in the rat corpus luteum during apoptosis-mediated luteolysis. Biol. Reprod. 60: 797–804
Cataldo NA, Dumesic DA, Goldsmith PC and Jaffe RB (2000) Immunolocalization of Fas and Fas ligand in the ovaries of women with polycystic ovary syndrome: relationship to apoptosis. Hum. Reprod. 15: 1889–1897
Kondo H, Maruo T, Peng X and Mochizuki M (1996) Immunological evidence for the expression of the Fas antigen in the infant and adult human ovary during follicular regression and atresia. J. Clin. Endocr. Metab. 81: 2702–2710
Adachi M, Suematsu S, Kondo T, Ogasawara J, Tanaka T, Yoshida N and Nagata S (1995) Targeted mutation in the Fas gene causes hyperplasia in peripheral lymphoid organs and liver. Nat. Genet. 11: 294–300
Tres LL and Kierszenbaum AL (1999) Cell death patterns of the rat spermatogonial cell progeny induced by sertoli cell geometric changes and Fas (CD95) agonist. Dev. Dynam. 214: 361–371
Koji T (2001) Male germ cell death in mouse testes: possible involvement of Fas and Fas ligand. Med. Electron Microsc. 34: 213–222
Francavilla S, D'Abrizio P, Cordeschi G, Pelliccione F, Necozione S, Ulisse S, Properzi, G. and Francavilla, F. (2002) Fas expression correlates with human germ cell degeneration in meiotic and post-meiotic arrest of spermatogenesis. Mol. Hum. Reprod. 8: 213–220
Lee J, Richburg JH, Younkin SC and Boekelheide K (1997) The Fas system is a key regulator of germ cell apoptosis in the testis. Endocrinology 138: 2081–2088
Koji T, Hishikawa Y, Ando H, Nakanishi Y and Kobayashi N (2001) Expression of Fas and Fas ligand in normal and ischemia–reperfusion testes: involvement of the Fas system in the induction of germ cell apoptosis in the damaged mouse testis. Biol. Reprod. 64: 946–954
Ogi S, Tanji N, Yokoyama M, Takeuchi M and Terada N (1998) Involvement of Fas in the apoptosis of mouse germ cells induced by experimental cryptorchidism. Urol. Res. 26: 17–21
Yin Y, DeWolf WC and Morgentaler A (1998) Experimental cryptorchidism induces testicular germ cell apoptosis by p53-dependent and -independent pathways in mice. Biol. Reprod. 58: 492–496
Peli J, Schroter M, Rudaz C, Hahne M, Meyer C, Reichmann E and Tschopp J (1999) Oncogenic Ras inhibits Fas ligand-mediated apoptosis by downregulating the expression of Fas. EMBO J. 18: 1824–1831
Koshimizu U, Nishioka H, Watanabe D, Dohmae K and Nishimune Y (1995) Characterization of a novel spermatogenic cell antigen specific for early stages of germ cells in mouse testis. Mol. Reprod. Dev. 40: 221–227
Kazama H and Yonehara S (2000) Oncogenic K-Ras and basic fibroblast growth factor prevent Fas-mediated apoptosis in fibroblasts through activation of mitogen-activated protein kinase. J Cell Biol. 148: 557–566
Kissel H, Timokhina I, Hardy MP, Rothschild G, Tajima Y, Soares V, Angeles M, Whitlow SR, Manova K and Besmer P (2000) Point mutation in kit receptor tyrosine kinase reveals essential roles for kit signaling in spermatogenesis and oogenesis without affecting other kit responses. EMBO J. 19: 1312–1326
Blume-Jensen P, Jiang G, Hyman R, Lee KF, O'Gorman S and Hunter T (2000) Kit/stem cell factor receptor-induced activation of phosphatidylinositol 3'-kinase is essential for male fertility. Nat. Genet. 24:157–162
Jordan SA, Speed RM and Jackson IJ (1999) Deficiency of Trp53 rescues the male fertility defects of Kit(W-v) mice but has no effect on the survival of melanocytes and mast cells. Dev. Biol. 215: 78–90
Yin Y, Stahl BC, DeWolf WC and Morgentaler A (2002) P53 and Fas are sequential mechanisms of testicular germ cell apoptosis. J. Androl. 23:64–70
Yoshida H, Takakura N, Kataoka H, Kunisada T, Okamura H and Nishikawa SI (1997) Stepwise requirement of c-kit tyrosine kinase in mouse ovarian follicle development. Dev. Biol. 184: 122–137
Eppig JJ, Schultz RM, O'Brien M and Chesnel F (1994) Relationship between the developmental programs controlling nuclear and cytoplasmic maturation of mouse oocytes. Dev. Biol. 164: 1–9
Inazawa Y and Yonehara S (1999) Fas-induced in vivo apoptosis in bone marrow: anti-Fas mAb-induced elimination and successive proliferation of Fas-expressing cells especially those of myeloid lineage. Cell Struct. Funct. 24: 151–159
Chen WS, Xu PZ, Gottlob K, Chen ML, Sokol K, Shiyanova T, Roninson I, Weng W, Suzuki R, Tobe K, Kadowaki T and Hay N (2001) Growth retardation and increased apoptosis in mice with homozygous disruption of the Akt1 gene. Genes. Dev. 15: 2203–2208
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS, Anderson MJ, Arden KC, Blenis J and Greenberg ME (1999) Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 96: 857–868
Panka DJ, Mano T, Suhara T, Walsh K and Mier JW (2001) Phosphatidylinositol 3-kinase/Akt activity regulates c-FLIP expression in tumor cells. J. Biol. Chem. 276: 6893–6896
Tanaka H, Pereira LA, Nozaki M, Tsuchida J, Sawada K, Mori H and Nishimune Y (1997) A germ cell-specific nuclear antigen recognized by a monoclonal antibody raised against mouse testicular germ cells. Int. J. Androl. 20: 361–366
Nishikawa S, Kusakabe M, Yoshinaga K, Ogawa M, Hayashi S, Kunisada T, Era T and Sakakura T (1991) In utero manipulation of coat color formation by a monoclonal anti-c-kit antibody: two distinct waves of c-kit-dependency during melanocyte development. EMBO J. 10: 2111–2118
Senju S, Negishi I, Motoyama N, Wang F, Nakayama KI, Nakayama K, Lucas PJ, Hatakeyama S, Zhang Q, Yonehara S and Loh DY (1996) Functional significance of the Fas molecule in naive lymphocytes. Int. Immunol. 8: 423–431
Lu XA and Werner D (1989) The complete cDNA sequence of mouse elongation factor 1 alpha (EF 1 alpha) mRNA. Nucleic Acids Res. 17: 442
Acknowledgements
We thank Drs. Ueda and Kanakura, Dr. Kikkawa and Dr. Nishikawa for providing mouse c-kit cDNA, caAkt cDNA and anti-c-kit mAb ACK2, respectively. We thank Drs. Hisahiro Yoshida and Shin-ichi Nishikawa for their useful advice. This study was supported in part by Grants-in-Aid from the Ministry of Education, Culture, Sports, Science and Technology of the Japanese Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by H Ichijo
Rights and permissions
About this article
Cite this article
Sakata, S., Sakamaki, K., Watanabe, K. et al. Involvement of death receptor Fas in germ cell degeneration in gonads of Kit-deficient Wv/Wv mutant mice. Cell Death Differ 10, 676–686 (2003). https://doi.org/10.1038/sj.cdd.4401215
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.cdd.4401215
Keywords
This article is cited by
-
XCI-escaping gene KDM5C contributes to ovarian development via downregulating miR-320a
Human Genetics (2017)
-
Mechanisms controlling germline cyst breakdown and primordial follicle formation
Cellular and Molecular Life Sciences (2017)
-
A survival Kit for pancreatic beta cells: stem cell factor and c-Kit receptor tyrosine kinase
Diabetologia (2015)
-
β-catenin/Tcf-signaling appears to establish the murine ovarian surface epithelium (OSE) and remains active in selected postnatal OSE cells
BMC Developmental Biology (2012)
