
Abstract
Telomerase reactivation and telomere maintenance are crucial in carcinogenesis and tumor progression. In this study, the relationships between telomere parameters, chromosomal instability and clinicopathological features were evaluated in hepatocellular carcinomas (HCCs). Telomere length (TL), telomerase activity (TA) and human telomerase reverse transcriptase (hTERT) mRNA levels were measured in 49 hepatitis B virus (HBV)-related HCCs and corresponding non-tumorous tissues. The results were compared with clinicopathological data, including differentiation, multipolar mitosis (MM), anaphase bridge, immunohistochemical stain results for cytokeratin 19 (CK19) and patient outcome. TL of HCCs ranged from 4.7 to 13.1 kb, and 44.4% of HCCs showed telomere lengthening. hTERT mRNA levels and TA were closely related (P=0.008), and were significantly higher in HCCs than non-tumorous tissues. TL was significantly higher in HCCs with strong TA (P=0.048), high hTERT mRNA levels (P=0.001) and poor differentiation (P=0.041). Frequent MM was associated with poor differentiation (P=0.007) and advanced stage (P<0.001). TA was positively correlated with MM, anaphase bridges and advanced stage (P=0.019, P=0.017 and P=0.029). Thirteen (28.3%) HCCs were CK19+ and demonstrated longer telomeres than CK19− HCCs (P=0.046). Overall survival was poor in HCCs with MM >0.4 per field (P=0.016), high TA (P=0.009) and high TL ratio (HCC/non-HCC) >0.8 (P=0.044). Our results show that long telomeres, high TA and high mitotic instability are poor prognostic markers for HBV-related HCCs and their close association suggests that telomere maintenance may be important for the progression of HCCs with high chromosomal instability to more aggressive ones.
Similar content being viewed by others

Main
The telomere, a distinctive DNA−protein structure at the end of chromosomes, stabilizes the natural end of the chromosome and protects against end-to-end fusion.1, 2 Telomerase is a ribonucleoprotein complex composed of two key components: a catalytic component (human telomerase reverse transcriptase, hTERT) and an RNA template. hTERT is a reverse transcriptase that synthesizes telomere repeats from the RNA template, and it is a limiting component in telomerase activity (TA).3, 4 During the early stages of carcinogenesis, cells undergo extensive proliferation until telomere length (TL) becomes critically shortened. Cells with dysfunctional telomeres gain further chromosomal instability through breakage–fusion–bridge cycles during replication, which have a catastrophic effect on cells. However, telomerase reactivation can stabilize the chromosome ends and produce aberrant, yet stable chromosomes; these cells then gain infinite life.5, 6 A current hypothesis is that telomere dysfunction leads to mitotic disturbances, giving rise to a high rate of chromatin bridges at anaphase and multipolar mitosis (MM).7 Tumors with high rates of anaphase bridging (such as ovarian, head and neck tumors) are characterized by a multimodal distribution of genomic imbalances, consistent with a dramatically increased rate of chromosomal rearrangement.8, 9
Hepatocellular carcinoma (HCC) is the seventh most common cancer worldwide, accounting for the highest number of adult malignancies in areas endemic for hepatitis B virus (HBV). Telomere shortening is an early event in multistep hepatocarcinogenesis, occurring in preneoplastic lesions of dysplastic nodules10, 11 and shortened telomeres have been reported to induce chromosomal instability in hepatocytes, especially important in HBV-related hepatocarcinogenesis.12, 13 However, the roles of telomere dysfunction and telomerase parameters in the progression of human HCCs have not been systematically analyzed.
In this study, the relationships between TL, TA, hTERT mRNA and pathological parameters (including differentiation, MM and anaphase bridge) and patient outcome were evaluated in HCCs and corresponding non-tumorous livers to clarify their roles in the progression of HCCs.
MATERIALS AND METHODS
Materials and Histopathological Examination
Forty-nine HCCs and their corresponding non-tumorous livers were retrieved from the archival files of Severance Hospital, Yonsei University College of Medicine. This study was approved by the Institutional Review Board of Severance Hospital, Yonsei University College of Medicine, and the specimens were supported by the Liver Cancer Specimen Bank from the National Research Resource Bank Program of the Korea Science and Engineering Foundation in the Ministry of Science and Technology. The tissues sampled from resected liver specimens were snap-frozen in liquid nitrogen and stored at −70°C. The patient population consisted of 36 males and 13 females, and their ages ranged from 35 to 69 years (53.7±9.18 years, mean±s.d.). All of the non-tumorous liver samples showed HBV-associated chronic hepatitis or cirrhosis. Representative sections were submitted for routine histological examination.
Hematoxylin and eosin (H&E)-stained tissue sections were examined under a light microscope using × 400 magnification for mitosis, MM and anaphase bridges. At least 10 high-power fields (HPFs) were reviewed. Metaphase and anaphase cells with chromosomes showing three or more spindle poles were classified as multipolar. Anaphase bridging was defined as an anaphase event in which more than two-thirds of the distance between anaphase poles was spanned by the bridging chromosome,14 to avoid the counting of lagging chromosomes.
Analysis of TL
Analysis was performed as described by a previous report.10 TL was measured from 2 μg HinfI-digested DNA. Hybridization was carried out with a 3′-end DIG-labeled d(TTAGGG)4 (Roche Molecular Biochemicals, Mannheim, Germany) at 37°C overnight. TL (telomere repeat frequency) was calculated using a previously described method;15 mean TL was obtained from two or three replicate experiments.
Telomeric Repeat Amplification Protocol Assay
TA was examined with a telomeric repeat amplification protocol (TRAP) using a TRAPeze® ELISA telomerase detection kit (Intergen, NY, USA).10 All samples were assayed with 0.2 μg lysate, providing activity in the linear range. Assays were duplicated; the mean values were taken as the TA of the samples. The activity was semiquantified by ELISA, as recommended by the manufacturer: high activity (+++): ≥1.5; moderate activity (++): 0.5–1.5; low activity (+): <0.5.
Real-Time Quantitative RT-PCR for hTERT mRNA
The analysis for hTERT mRNA was performed using real-time quantitative RT-PCR, based on TaqMan methodology and using an ABI PRISM 7700 sequence detection system (Perkin-Elmer, CA, USA). A standard RNA, prepared by in vitro run-off transcription with T7 RNA polymerase, was used for a standard curve. This was prepared from 5 to 6 dilutions of the standard hTERT RNA (3 × 101 to 3 × 105 copies) and GAPDH RNA (3 × 102 to 3 × 106 copies). PCR and thermal cycling were carried out as described in a previous report.16 Sequences of the hTERT primers and probe were as follows: hTERT-F, 5′-CACGCGAAAACCTTCCTCA-3′; hTERT-R, 5′-CAAGTTCACCACGCAGCC-3′, and the probe, 5′-(FAM)-CTCAGGGACACCTCGGACCAGGGT-(TAMRA)-3′. Sequences of the GAPDH primers and probes were as follows: GAPDH-F, 5′-CCTCTGACTTCAACAGCGACAC-3′; GAPDH-R, 5′-GTGGTCGTTGAGGGCAATG-3′; and the probe, 5′-(FAM)-CACTCCTCCACCTTTGACGCTGGG-(TAMRA)-3′.
Immunohistochemistry for Cytokeratin 19
Paraffin sections were deparaffinized with xylene and rehydrated with graded alcohol. After washing in distilled water the sections were immersed in 3% hydrogen peroxide to block endogenous peroxidases. Antigen retrieval was performed by boiling the sections in 100 mM sodium citrate (pH 6.0) for 15 min in a microwave oven. Monoclonal primary antibody (cytokeratin 19, DAKO, Glostrup, Denmark) at a 1:50 dilution was applied for 30 min at room temperature followed by washing in PBS. Incubation with the secondary antibody was carried out using the DAKO EnVision Rabbit/Mouse kit for 30 min at room temperature, developed with diaminobenzidine (DAKO), and counterstained with hematoxylin. Strong cytoplasmic staining in at least more than 5% of the tumor cells classified HCCs as CK19+.
Statistical Analysis
Statistical analysis was performed using the SPSS software package, version 12.0 (SPSS Inc., Chicago, IL, USA). Differences in telomere parameters among various groups of patients discriminated for histopathologic parameters and CK19 expression status were analyzed by the Kruskal–Wallis and Mann–Whitney two-sample tests. Correlations between histopathological parameters and telomere parameters were analyzed by the Spearman's correlation test. The Kaplan–Meier method was used for analysis of overall survival. A P-value of less than 0.05 was considered statistically significant.
RESULTS
Histopathological Features and Mitotic Instability in Hepatocarcinogenesis
The HCCs ranged from 1 to 18 cm in size (5.7±4.1 cm). According to Edmondson's grading scores, the differentiation of HCCs was as follows: grade I in 2 cases, grade II in 24 cases, grade III in 22 cases and grade IV in 1 case. The TNM stages of the HCCs according to the 2002 American Joint Committee on Cancer (AJCC)/International Union Against Cancer (UICC) system were as follows: stage I in 17 cases, stage II in 16 cases, stage III in 10 cases (stage IIIA in 8 cases, stage IIIB in 1 case and stage IIIC in 1 case) and stage IV in 6 cases. The TNM stage and histologic grade of the HCCs were significantly correlated (P=0.037). The mitotic count of HCCs ranged from 0 to 17 mitotic figures per HPF (3.09–3.22), and a significant positive correlation was shown between mitotic count and differentiation (P=0.033) and TNM stage (P<0.001). Clinicopathological characteristics are summarized in Table 1.
Anaphase bridges were found in 15 HCCs (30.6%; 0.03±0.06 per HPF) (Figure 1a). MM were found in 35 cases (71.4%), and the number of MM per HPF ranged from 0 to 3.9 (0.58±0.917) (Figure 1b). Anaphase bridges and MM were significantly positively correlated (P=0.001). MM were more frequent in poorly differentiated HCCs (P=0.007) and in HCCs of advanced stage (P<0.001), while anaphase bridges were not significantly correlated with histological parameters.

Hepatocellular carcinoma showing anaphase bridging (arrow) (a) and multipolar mitosis (arrows) (b) (hematoxylin and eosin, original magnification × 400).
Relationships between Telomere Parameters in Hepatocarcinogenesis
TLs were measured in 45 HCCs and compared with their adjacent non-neoplastic tissues (Table 1). TLs ranged from 4.7 to 13.1 kb in HCCs and from 6.3 to 12.6 kb in non-HCCs and the TL ratios of HCCs to non-tumorous livers (TLR) ranged from 0.61 to 1.54 (0.97±0.204). Twenty-five HCCs (55.6%) demonstrated shorter telomeres than their adjacent non-tumorous livers (7.2±1.59 and 8.7±1.69 kb, respectively), while the remaining 20 HCCs (44.4%) appeared to have longer telomeres than non-neoplastic tissues (9.4±1.23 and 8.3±0.94 kb, respectively).
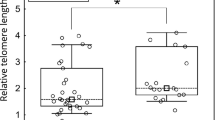
TA was measured in 40 HCCs and their adjacent non-neoplastic tissues (Table 1) and the data were partly reported earlier.16 TL in HCCs with high TA were significantly higher than in those with low or moderate TA (8.8±2.10 and 7.7±1.62, respectively; P=0.048) (Figure 2). There was also a significant positive correlation between TLR and TA (P=0.038).

Telomere length according to telomerase activity (TA) in hepatocellular carcinomas (HCCs) (mean±s.d.). Telomere length is significantly higher in HCCs with strong TA than HCCs with low or moderate TA (P<0.05) (TRF(T), telomere repeat frequency of tumor).
hTERT mRNA levels were determined in the same tissues used for the determination of TA, and ranged from 15 to 28115 (1477.5±4483.30) in HCCs and from 0 to 166 (38.6±49.39) in non-neoplastic tissues (Table 1). In all cases, the HCCs demonstrated higher levels of hTERT mRNA than adjacent non-tumorous livers. A significant positive correlation between hTERT mRNA level and TA was found in the HCCs (P=0.008). In addition, higher hTERT mRNA levels were significantly associated with longer telomeres (P=0.001) (Figure 3a) and higher TLR (P=0.034).

The relationships between hTERT mRNA levels and telomere length (a), mitosis (b) and multipolar mitosis (c) (hTERT(T), hTERT mRNA levels of the tumor; M/f, mitotic figures per × 400 field; MM/f, multipolar mitosis per × 400 field). Significant positive correlations are seen between hTERT mRNA levels and telomere length (P=0.001), mitosis (P<0.001) and multipolar mitosis (P=0.011).
Relationship between Telomere Parameters, Histopathological Parameters and Mitotic Instability
TLs were longer in HCCs with poorer differentiation (P=0.041; 7.7±1.80 kb in grade I and II HCCs vs 8.7±1.66 kb in grade III and IV HCCs). TL showed positive correlations with mitotic activity (P=0.002).
High TA was significantly associated with advanced tumor stage, high mitotic rates, frequent MM and anaphase bridges (P=0.029, P=0.032, P=0.019 and P=0.017, respectively). hTERT mRNA levels were positively correlated with advanced tumor stage, mitotic count and MM (P=0.012, P<0.001 and P=0.011, respectively) (Figure 3b and c), but no significant relations were seen between hTERT mRNA levels and anaphase bridges.
Relationships between CK19 Positivity, Telomere Parameters and Mitotic Instability
Immunohistochemical stain for the biliary epithelial cell marker CK19 was performed in 46 HCCs, and 13 (28.3%) cases showed strong cytoplasmic staining for CK19 in more than 5% of the tumor cells (Table 1). In fact, 5 of the 13 CK19+ cases focally demonstrated histological features of progenitor cell or biliary differentiation: areas of relatively small and monomorphous tumor cells with scant cytoplasm, partly arranged in tubule-like structures. Interestingly, CK19+ HCCs demonstrated significantly longer telomeres than those with CK19 negativity (8.97±1.34 vs 7.89±1.87 kb; P=0.046) (Figure 4) and showed increased hTERT mRNA levels compared to CK19− HCCs (although not statistically significant).

Telomere length according cytokeratin 19 expression status in HCCs (mean±s.d.). Telomeres of CK19+ HCCs were significantly longer than those of CK19− tumors (P<0.05).
Effects of Telomere Parameters and Mitotic Instability on Prognosis
Follow-up visits were possible in 45 of the 49 cases, with the follow-up period ranging from 4 to 42 months (mean=22 months). Local recurrences were seen in 19 of the 49 patients (38.8%) after a mean of 14 months of disease-free survival. Eleven patients (24.4%) died of hepatic failure or metastasis 4 to 24 months following operation (Table 1).
A poorer prognosis was seen in HCCs with an MM ≥0.4 per HPF (P=0.016). Overall survival was poor when the TLR was greater than 0.8 (P=0.044) and in cases with high TA (P=0.009) (Figure 5a–c). When combining TLR with TA, three prognostic groups could be defined (Figure 5d). Group 1 comprised HCCs with shortened telomeres (TLR <0.8) regardless of TA and demonstrated a survival rate of 100%. Group 2 consisted of HCCs with TLR ≥0.8 and low/moderate TA (survival rate: 80%), and group 3 HCCs were those with TLR ≥0.8 and high TA (survival rate: 54.5%). The differences in survival between the three groups were statistically significant (group 1 vs group 3: P=0.037, group 2 vs group 3: P=0.043), demonstrating that the combination of maintained or lengthened telomeres and high TA serve as ominous prognostic factors in HCCs. CK19 expression status of HCCs demonstrated no significant influence on patient survival.

Kaplan–Meier curve for overall survival of HCC patients according to telomerase activity (TA) (a), telomere length ratio (TL ratio) of the HCC to the non-tumorous livers (b), and multipolar mitosis (MM) count (c). Three prognostic groups could be defined after combination of TL ratio with TA (d) group 1, TL ratio <0.8 regardless of TA status; group 2, TL ratio ≥0.8 and low/moderate TA; group 3, TL ratio ≥0.8 and high TA.
DISCUSSION
Telomere dysfunction plays an important role in carcinogenesis by the induction of chromosomal instability through the breakage–fusion–bridge cycle of affected cells, and telomere shortening has been shown to be crucial in the initiation of carcinogenesis.17, 18 Previous studies on dysplastic nodules and dysplastic foci of the liver demonstrated that telomere shortening occurs in the early stages of hepatocarcinogenesis,10, 11, 19 again providing evidence for the importance of telomere dysfunction in cancer initiation. However, not all preneoplastic lesions are considered to be capable of fully advancing to cancer. Only those cells that find a way to maintain their telomeres, thus allowing unlimited cell division and some degree of chromosomal instability, will be able to progress to more advanced malignant tumors. Therefore, while telomere shortening and chromosomal instability take place in the earlier stages of carcinogenesis, cancer progression is associated with telomere maintenance. In this study, we analyzed TLs of HBV-related HCCs, and found that although just over a half (55.6%) of our HCCs had shorter telomeres compared to non-neoplastic livers, a substantial percentage (44.4%) demonstrated unchanged or even increased TLs. In addition, TL was positively correlated with poorer differentiation. Nakajima et al20 reported that HCCs of less than 5 cm showed a negative correlation between TL and tumor size, that is, TLs decreased with increasing tumor size, whereas tumors larger than 5 cm showed size-independent TLs and were associated with highly malignant clinicopathological features. As TLs of HCCs and corresponding non-neoplastic livers vary for each individual, the TLR was calculated in each case as in previous studies21, 22 to analyze the alteration status of TL relative to the paired non-tumorous tissue. Interestingly, we found that the group of patients with TLR ≥0.8 showed a poorer overall survival, implying that maintenance or elongation of telomeres in HCCs may serve as a poor prognostic factor. The association of long telomeres with advanced stages and poorer survival has also been recently reported in other tumors including colorectal cancers and head and neck tumors.21, 22, 23, 24, 25, 26
In this study, all HCCs demonstrated higher hTERT mRNA levels than their corresponding non-tumorous tissues, and the positive correlation between hTERT level and TA in our study supports previous studies which showed that hTERT is a key regulator of TA in hepatocarcinogenesis.27, 28, 29, 30 In addition, hTERT mRNA levels were positively correlated with TLs and HCCs with high TA showed minimal shortening or even lengthening of telomeres, suggesting that telomerase may play a significant role in telomere maintenance or lengthening during HCC progression. Furthermore, advanced tumor stage and poorer overall survival were seen in HCCs with strong TA supporting previous reports that showed HCCs with higher TA were more aggressive with frequent recurrence after operation.31, 32, 33 In our study, the difference in overall survival according to hTERT mRNA levels failed to reach statistical significance, unlike previous studies which showed that high hTERT expression was associated with a poorer prognosis in colorectal carcinomas and ependymomas.34, 35 However, we found that high hTERT mRNA levels were significantly correlated with more advanced tumor stage.
Chromosomal instability has been reported to play an important role in the pathways of HBV-related hepatocarcinogenesis and in HCCs associated with poor differentiation or poor prognosis.12, 13 In this study, MM and anaphase bridges were demonstrated in 71.4 and 30.6% of HBV-related HCCs, respectively, and the two features were significantly positively correlated. MM were more prevalent in poorly differentiated HCCs and in those with advanced stage, and HCCs with MM ≥0.4 per HPF showed a poor overall survival in this study. To some extent, these results are consistent with a previous report which demonstrated a close correlation between the morphological dedifferentiation of HCCs and an increase in chromosomal heterogeneity.36
In an animal study of successive generations of telomerase-deficient mice, short dysfunctional telomere was reported to be associated with a significant increase in the number of early neoplastic lesions and a reciprocal decline in the occurrence of high-grade malignancies.37 Similarly, hTERT-mediated telomere stabilization is suggested to be critical for the progression and prognosis of colorectal cancers.23 Lechel et al38 demonstrated in their study of p53-mutant mice that telomerase is a critical component for in vivo progression of p53-mutant HCC with short telomeres in chronically damaged liver—telomerase limits the accumulation of telomere dysfunction and chromosomal instability—and, therefore, telomerase deletion limits the progression of p53-mutant HCC with short telomeres. In our HCCs, those with chromosomal instability (ie frequent MM and anaphase bridges) showed significantly higher TA and hTERT mRNA levels, again implying that these genetically instable tumors require telomerase to prevent complete genomic deterioration and to progress to more aggressive and advanced HCCs.
The concept of stem cell carcinogenesis has received a lot of attention lately. Most direct evidence for the hypothesis of cancer stem cells has come from studies on hematologic malignancies. Shortening of telomeres has been shown in lymphoid and myeloid leukemias suggesting that telomere loss may contribute to cancer stem cell initiation; however, cancer stem cells may acquire immortality by mutational events in telomere-lengthening mechanisms such as telomerase activation, resulting in maintenance of TL, disease progression, poor prognosis and increased risk of recurrence in treated patients.39, 40, 41 Stem cells have also been implicated in hepatocarcinogenesis; several studies have demonstrated that as much as 28–50% of human HCCs express markers of progenitor/biliary cells such as CK7, CK19 and OV6, and also markers of hematopoietic stem cells such as c-kit and CD34.42, 43, 44 These HCCs have been characterized by an immature phenotype intermediate between biliary epithelial cells and hepatocytes, and CK19 expression in HCC has been associated with a worse prognosis.45, 46, 47, 48 It is worth noting from our study that telomeres of CK19+ HCCs were significantly longer than those that were CK19−, which could support previous reports that HCCs with progenitor cell features have a poor prognosis. However, no definite association between CK19 expression status and prognosis was found in our study, probably attributed to the paucity of CK19+ cases. Therefore, a more extensive study will be necessary for a further understanding of the relationship between hepatic stem cell carcinogenesis and telomere dynamics.
In conclusion, TL, TA and mitotic instability are useful prognostic markers for HBV-related HCCs. The close association between TL, TA, mitotic instability and poor prognosis suggests that maintenance of telomere may be important for the progression of HCCs with high chromosomal instability to more aggressive ones. Telomerase inhibition and telomere regulation could be a possible target in the treatment of both early and advanced HCCs.
References
Collins K . Mammalian telomeres and telomerase. Curr Opin Cell Biol 2000;12:378–383.
Maser RS, DePinho RA . Connecting chromosomes, crisis, and cancer. Science 2002;297:565–569.
Nakayama J, Tahara H, Tahara E, et al. Telomerase activation by hTERT in human normal fibroblasts and hepatocellular carcinomas. Nat Genet 1998;18:65–68.
Nakamura TM, Morin GB, Chapman KB, et al. Telomerase catalytic subunit homologs from fission yeast and human. Science 1997;277:955–959.
Ishikawa F . Telomere crisis, the driving force in cancer cell evolution. Biochem Biophys Res Commun 1997;230:1–6.
Murnane JP, Sabatier L . Chromosome rearrangements resulting from telomere dysfunction and their role in cancer. Bioessays 2004;26:1164–1174.
Gisselsson D, Hoglund M . Connecting mitotic instability and chromosome aberrations in cancer-can telomeres bridge the gap? Semin Cancer Biol 2005;15:13–23.
Gisselsson D, Jonson T, Yu C . Centrosomal abnormalities, multipolar mitosis, and chromosomal instability in head and neck tumours with dysfunctional telomeres. Br J Cancer 2002;87:202–207.
Gisselsson D, Lv M, Tsao SW, et al. Telomere-mediated mitotic disturbance in immortalized ovarian epithelial cells reproduce chromosomal losses and breakpoints from ovarian carcinoma. Gene Chromosomes Cancer 2005;42:22–33.
Oh BK, Chae KJ, Park C, et al. Telomere shortening and telomerase reactivation in dysplastic nodules of human hepatocarcinogenesis. J Hepatol 2003;39:786–792.
Oh BK, Kim YJ, Park C, et al. Up-regulation of telomere-binding proteins, TRF1, TRF2, and TIN2 is related to telomere shortening during human multistep hepatocarcinogenesis. Am J Pathol 2005;166:73–80.
Laurent-Puig P, Legoix P, Bluteau O, et al. Genetic alterations associated with hepatocellular carcinomas define distinct pathways of hepatocarcinogenesis. Gastroenterology 2001;120:1763–1773.
Plentz RR, Caselitz M, Bleck JS, et al. Hepatocellular telomere shortening correlates with chromosomal instability and the development of human hepatocellular carcinoma. Hepatology 2004;40:80–86.
Rudolph KL, Millard M, Bosenberg MW, et al. Telomere dysfunction and evolution of intestinal carcinoma in mice and humans. Nat Genet 2001;28:155–159.
Kruk PA, Rampino NJ, Bohr VA . DNA damage and repair in telomeres: relation to aging. Proc Natl Acad Sci USA 1995;92: 258–262.
Oh BK, Chae KJ, Park C, et al. Molecular analysis of PinX1 in human hepatocellular carcinoma. Oncol Rep 2004;12:861–866.
Meeker AK, Hicks JL, Iacobuzio-Donahue CA, et al. Telomere length abnormalities occur early in the initiation of epithelial carcinogenesis. Clin Cancer Res 2004;10:3317–3326.
Gisselsson D, Jonson T, Petersen A, et al. Telomere dysfunction triggers extensive DNA fragmentation and evolution of complex chromosome abnormalities in human malignant tumors. Proc Natl Acad Sci USA 2001;98:12683–12688.
Plentz RR, Park YN, Lechel A, et al. Telomere shortening and inactivation of cell cycle checkpoints characterize human hepatocarcinogenesis. Hepatology 2007;45:968–976.
Nakajima T, Katagishi T, Moriguchi M, et al. Tumor size-independence of telomere length indicates an aggressive feature of HCC. Biochem Biophys Res Commun 2004;325:1131–1135.
Gertler R, Rosenberg R, Stricker D, et al. Telomere length and human telomerase reverse transcriptase expression as markers for progression and prognosis of colorectal carcinoma. J Clin Oncol 2004;22:1807–1814.
Garcia-Aranda C, de Juan C, Diaz-Lopez A, et al. Correlations of telomere length, telomerase activity, and telomeric-repeat binding factor 1 expression in colorectal carcinoma: prognostic indications. Cancer 2006;106:541–551.
Engelhardt M, Drullinsky P, Guillem J, et al. Telomerase and telomere length in the development and progression of premalignant lesions to colorectal cancer. Clin Cancer Res 1997;3:1931–1941.
Patel MM, Parekh LJ, Jha FP, et al. Clinical usefulness of telomerase activation and telomere length in head and neck cancer. Head Neck 2002;24:1060–1067.
Bisoffi M, Heaphy CM, Griffith JK . Telomeres: prognostic markers for solid tumors. Int J Cancer 2006;119:2255–2260.
Ohali A, Avigad S, Ash S, et al. Telomere length is a prognostic factor in neuroblastoma. Cancer 2006;107:1391–1399.
Nagao K, Tomimatsu M, Endo H, et al. Telomerase reverse transcriptase mRNA expression and telomerase activity in hepatocellular carcinoma. J Gastroenterol 1999;34:83–87.
Hisatomi H, Nagao K, Kanamaru T, et al. Levels of telomerase catalytic subunit mRNA as a predictor of potential malignancy. Int J Oncol 1999;14:727–732.
Takahashi S, Kitamoto M, Takaishi H, et al. Expression of telomerase component genes in hepatocellular carcinomas. Eur J Cancer 2000;36:496–502.
Saretzki G, Petersen S, Petersen I, et al. hTERT gene dosage correlates with telomerase activity in human lung cancer cell lines. Cancer Lett 2002;176:81–91.
Suda T, Isokawa O, Aoyagi Y, et al. Quantitation of telomerase activity in hepatocellular carcinoma: a possible aid for a prediction of recurrent diseases in the remnant liver. Hepatology 1998;27:402–406.
Kobayashi T, Kubota K, Takayama T, et al. Telomerase activity as a predictive marker for recurrence of hepatocellular carcinoma after hepatectomy. Am J Surg 2001;181:284–288.
Kishimoto K, Fujimoto J, Takeuchi M, et al. Telomerase activity in hepatocellular carcinoma and adjacent liver tissues. J Surg Oncol 1998;69:119–124.
Gertler R, Rosenberg R, Stricker D, et al. Prognostic potential of the telomerase subunit human telomerase reverse transcriptase in tumor tissue and nontumorous mucosa from patients with colorectal carcinoma. Cancer 2002;95:2103–2111.
Tabori U, Ma J, Carter M, et al. Human telomere reverse transcriptase expression predicts progression and survival and pediatric intracranial ependymoma. J Clin Oncol 2006;24:1522–1528.
Wilkens L, Flemming P, Gebel M, et al. Induction of aneuploidy by increasing chromosomal instability during dedifferentiation of HCC. Proc Natl Acad Sci USA 2004;101:1309–1314.
Farazi PA, Glickman J, Jiang S, et al. Differential impact of telomere dysfunction on initiation and progression of hepatocellular carcinoma. Cancer Res 2003;63:5021–5027.
Lechel A, Holstege H, Begus Y, et al. Telomerase deletion limits progression of p53-mutant hepatocellular carcinoma with short telomeres in chronic liver disease. Gastroenterology 2007;132:1465–1475.
Ohyashiki JH, Sashida G, Tauchi T, et al. Telomeres and telomerase in hematologic neoplasia. Oncogene 2002;21:680–687.
Ju Z, Rudolph KL . Telomeres and telomerase in cancer stem cells. Eur J Cancer 2006;42:1197–1203.
Hiyama E, Hiyama K . Telomere and telomerase in stem cells. Br J Cancer 2007;96:1020–1024.
Roskams T . Liver stem cells and their implication in hepatocellular and cholangiocarcinoma. Oncogene 2006;25:3818–3822.
Wu PC, Lai VC, Fang JW, et al. Hepatocellular carcinoma expressing both hepatocellular and biliary markers also expresses cytokeratin 14, a marker of bipotential progenitor cells. J Hepatol 1999;31:965–966.
Kim H, Park C, Han KH, et al. Primary liver carcinoma of intermediate (hepatocyte–cholangiocyte) phenotype. J Hepatol 2004;40:298–304.
Durnez A, Verslype C, Nevens F, et al. The clinicopathological and prognostic relevance of cytokeratin 7 and 19 expression in hepatocellular carcinoma. A possible progenitor cell origin. Histopathology 2006;49:138–151.
Lee J-S, Heo J, Libbrecht L, et al. A novel prognostic subtype of human hepatocellular carcinoma derived from hepatic progenitor cells. Nat Med 2006;12:410–416.
Uenishi T, Kubo S, Yamamoto T, et al. Cytokeratin 19 expression in hepatocellular carcinoma predicts early postoperative recurrence. Cancer Sci 2003;94:851–857.
Yoon DS, Jeong J, Park YN, et al. Expression of biliary antigen and its clinical significance in hepatocellular carcinoma. Yonsei Med J 1999;40:472–477.
Acknowledgements
This work was supported by a Grant (0620210) from the National R&D Program for Cancer Control, Ministry of Health & Welfare, Republic of Korea, a faculty grant from Yonsei University College of Medicine for 2007, and partly by KOSEF through the Cancer Metastasis Research Center at Yonsei University College of Medicine (R11-2000-082-02008-0) (to B-K Oh).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Oh, BK., Kim, H., Park, Y. et al. High telomerase activity and long telomeres in advanced hepatocellular carcinomas with poor prognosis. Lab Invest 88, 144–152 (2008). https://doi.org/10.1038/labinvest.3700710
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/labinvest.3700710
Keywords
This article is cited by
-
A telomerase regulation-related lncRNA signature predicts prognosis and immunotherapy response for gastric cancer
Journal of Cancer Research and Clinical Oncology (2023)
-
The role of telomeres and telomerase in cirrhosis and liver cancer
Nature Reviews Gastroenterology & Hepatology (2019)
-
Frequency and geographic distribution of TERT promoter mutations in primary hepatocellular carcinoma
Infectious Agents and Cancer (2017)
-
Association between telomere length and survival in cancer patients: a meta-analysis and review of literature
Frontiers of Medicine (2016)
-
Alterations of telomere length and mtDNA copy number are associated with overall survival in hepatocellular carcinoma patients treated with transarterial chemoembolization
Cancer Chemotherapy and Pharmacology (2016)
