
Abstract
Alcohol withdrawal symptoms, particularly negative emotional states, can persist for months following the removal of alcohol. These protracted withdrawal symptoms have been implicated as an important trigger of relapse to excessive drinking in alcoholics and may represent a long lasting shift in affective tone as a result of chronic alcohol exposure. It was shown previously that ethanol-dependent rats increased their operant responding for ethanol when tested during the first 12 hr after withdrawal. The purpose of the present experiments was to determine the persistence of this finding by examining operant oral ethanol self-administration in rats with a history of physical dependence upon ethanol, detoxified and then allowed a two week period of protracted abstinence. The results of these experiments indicate that operant responding for ethanol was enhanced during protracted abstinence by 30–100% and remained elevated for 4–8 weeks post acute withdrawal. These results have important implications for understanding the characteristics and mechanisms underlying vulnerability to relapse.
Similar content being viewed by others

Main
Excessive drinking of ethanol in animals can be produced by a number of factors including altering palatability, genetics, and history of consumption. There is evidence that certain symptoms of withdrawal can persist for a number of weeks or even months following chronic ethanol exposure in humans (Kissin 1979; Begleiter and Porjesz 1979; Alling et al. 1982; Roelofs 1985; Grant et al. 1987) as well as in animals (Begleiter and Porjesz 1979). In human alcoholics, one of the factors leading to excessive drinking is the use of alcohol to relieve or avoid withdrawal symptoms (U.S. Department of Health and Human Services 1990). Fatigue and tension persisted for approximately five weeks following withdrawal in a group of 68 chronic alcoholics (Alling et al. 1982), and periods of hyperventilatory symptomology and anxiety (as determined by the Spielberger State-Trait Anxiety Inventory, a self-report inventory), which correlated with intensity of alcohol craving, were reported for up to nine months following withdrawal in a group of 37 chronic alcoholics (Roelofs 1985).
In a larger cohort of 312 abstinent alcoholics, 20–25% of them showed signs of anxiety and depression, as determined from the Symptom Check-List 90 (self-report inventory with coverage of areas of symptomology and psychopathology) six months to two years following withdrawal (De Soto et al. 1985). In a follow-up study, it was shown that distress-related symptoms correlated with relapse in alcoholics who were abstinent for less than two years (De Soto et al. 1989). The more protracted symptoms tended to be subacute, were often affective in nature, and appeared to precede relapses into uncontrolled alcohol drinking. For example, depression and anxiety associated with withdrawal were found to provoke drinking in 83 of 100 male alcoholics who experienced these symptoms (Hershon 1977). These patients reported that they drank alcoholic beverages when they experienced anxiety and depressed mood. In another study, both male and female alcoholics reported negative emotions as the most common trigger of relapse (Annis et al. 1998). The clinical literature suggests that alterations in affective state persist for quite some time following alcohol withdrawal and may actually be partly responsible for some relapse episodes in alcoholics.
Potential rodent models of excessive ethanol drinking include the alcohol deprivation effect in nondependent animals, ethanol self-administration in dependent withdrawing animals, and ethanol self-administration in animals with a history of dependence following periods of abstinence. The alcohol deprivation effect is a transient increase in ethanol self-administration in animals following periods of abstinence from ethanol for several days to several weeks (Sinclair and Senter 1967; Spanagel et al. 1996; Heyser et al. 1997; Hölter et al. 1998). This increase in ethanol intake following periods of abstinence has been observed in animals having an extended history of ethanol drinking, but which did not necessarily show evidence of physical dependence on ethanol (i.e., no evidence of overt physical withdrawal symptoms).
Animal studies directed at examining the effects of withdrawal on ethanol self-administration have produced varying results (Myers et al 1972; Deutsch and Koopmans 1973; Samson and Falk 1974; Deutsch and Walton 1977; Begleiter 1975; Cappell and LeBlanc 1981; Winger 1988; Edwards 1990; Schulteis et al 1996), but more recent work has shown that there are two factors which appear to increase the likelihood of ethanol drinking in response to withdrawal. First, ethanol must be established as a reinforcer prior to dependence induction (Meisch 1983) and second, the subjects must learn an association between ethanol drinking and alleviation of withdrawal symptoms (Meisch 1994). In rats trained to self-administer ethanol prior to the induction of dependence and then tested during repeated withdrawal sessions, responding for ethanol was enhanced during withdrawal (Roberts et al. 1996). These results suggested that the rats learned across repeated withdrawal sessions to minimize withdrawal symptomology by self-administering ethanol. In addition, there have been a few studies examining ethanol self-administration in animals made dependent on ethanol and then allowed longer periods of abstinence. Denoble and Begleiter (1978) found that monkeys which received infusions of 5 g/kg ethanol every other day for 20 days showed increased intravenous ethanol self-administration relative to sucrose-intubated controls when tested four months later. In another study, rats consumed 37% of ethanol in a two bottle-choice procedure (ethanol vs. water) following at least two months of exposure to ethanol as the sole liquid (Ahlenius and Engel 1974), a percentage much higher than would be expected from ethanol-naive rats of this strain. In a recent study, the breaking points (maximum number of lever presses the animal performs to obtain a single ethanol delivery) of rats exposed to an ethanol liquid diet were higher than control rats over several days following withdrawal as well as 2–3 weeks following the third withdrawal (Brown et al. 1998). These data suggest that the excessive ethanol self-administration observed during withdrawal may persist for longer than any physical withdrawal symptoms.
Allostasis is a form of physiological regulation first hypothesized to describe the fluctuations in blood pressure and immune system function that are not well explained by homeostasis (Sterling and Eyer 1988). Homeostasis is the holding constant of internal parameters within the normal range. However, allostasis is a maintenance of stability at any level outside the normal range and is achieved by varying the internal milieu to match perceived and anticipated environmental demands. Allostasis as a form of regulation allows for the continuous reevaluation and readjustment of all physiological parameters towards new needs as well as for anticipation of such needs, and thus, presumably involves the brain's control over physiological systems. When demands on an individual are chronic, the set point for functioning is altered and may be maintained at such an altered point indefinitely. Although this altered set point may appear appropriate to the conditions, it may be in the pathological range in that any additional perturbation can produce dysregulation. The transition from controlled to excessive drug intake has recently been hypothesized to involve such a change in an internal reference mechanism or set point (Koob and Le Moal 1997; Ahmed and Koob 1998). Chronic drug or alcohol exposure may elicit allostasis within the brain's reward mechanisms as a means to maintain stability in the face of chronic demand. This allostatic change in set point has been hypothesized to be long-lasting and to play a role in the vulnerability to relapse (Koob and Le Moal 1997).
The persistent affective withdrawal symptoms observed in human alcoholics also may be a reflection of a shift in reward set point such that reward system functioning is decreased below baseline in the drug-free state. It has been argued that global allostatic changes associated with chronic demands lead to an unpleasant withdrawal state when the demands are lifted and provides the basis for seeking conditions of high demand (Sterling and Eyer 1988). This would explain vulnerability to relapse as a means to reachieve a normal reward state and the escalated ethanol intake as an attempt to re-experience the same level of rewarding effects previously experienced with ethanol. Examining the mechanisms responsible for protracted abstinence and its role in further ethanol consumption will be important in understanding the allostatic alterations, or alterations in reward thresholds, which may be responsible for the chronically relapsing nature of ethanol addiction.
The purpose of the current experiments was to examine potential alterations in operant oral ethanol self-administration following protracted abstinence in rats with a history of dependence. Based on the conceptualization of ethanol addiction as reflecting an enhancement in drug reward set point or threshold (Koob and Le Moal 1997), it was hypothesized that rats with a history of dependence would show a long lasting increase in baseline ethanol responding. Therefore, the model described herein has the potential to provide a means to examine long lasting alterations in the mechanisms mediating ethanol reward which may be partly responsible for the chronically relapsing nature of alcoholism in humans. A paradigm was used in which rats were first trained to lever press for ethanol and then made dependent on ethanol in vapor chambers (Roberts et al. 1996). Thus, ethanol was established as a reinforcer prior to the forced induction of dependence using the ethanol vapor inhalation approach. In one group, rats were allowed to operantly respond for ethanol during early withdrawal and then tested following a period of protracted abstinence. Rats in another group were not allowed to respond for ethanol during early withdrawal prior to testing, following protracted abstinence. In addition, the alcohol deprivation effect was examined following periods of abstinence in rats with or without a history of physical dependence. For the purposes of this manuscript, early withdrawal is defined as 0–12 hr following removal from chronic ethanol exposure, whereas protracted withdrawal or protracted abstinence refers to two or more weeks following removal from chronic ethanol exposure.
MATERIALS AND METHODS
General Methods
Animals
Male Wistar rats obtained from Charles River Laboratory (Kingston, NY) were used in Experiments 1–3 (n = 48), whereas Wistar rats originally derived from Charles River Laboratory (Kingston, NY), but bred in the Beckman Laboratories of The Scripps Research Institute, were used in Experiment 4 (n = 12). These latter Wistar rats are bred using a circular-pair random system of breeding in order to maintain genetic heterogeneity and new breeders are obtained from Charles River as determined by our internal Genetics Advisory Board. Separate groups of rats were used for each of the four experiments. Body weight was 180–200 g at the start of the experiments. Rats were housed 2–3 per cage with food and water available ad libitum, except for three days of water restriction at the initiation of operant testing (see below). Lights were on a 12-hr light/dark cycle, with lights on at 6:00 AM. All procedures met the guidelines of the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Operant Ethanol Self-Administration
Ethanol dilutions (5, 8, and 10% w/v) were prepared with 95% ethyl alcohol and water. Saccharin (Sigma Chemical Co., St. Louis, MO) was added to water or the ethanol solutions to achieve a concentration of 0.2% w/v.
Standard operant chambers (Coulbourn Instruments, Allentown, PA) housed in sound-attenuated, ventilated cubicles were used for ethanol self-administration. Syringe pumps dispensed ethanol and water into two stainless steel drinking cups mounted 4 cm above the grid floor in the middle of one side panel. Two retractable levers were located 4.5 cm to either side of the drinking cups. Fluid delivery and recording of operant responding were controlled by microcomputer. A continuous reinforcement (FR1) schedule was used, and responding resulted in delivery of 0.1 ml of fluid. During the 0.5 second in which the pumps are activated, lever presses are not recorded.
Rats were trained to lever press for ethanol using an adaptation of Samson's sweetened solution fading procedure (Samson 1986). The details of this procedure have been described elsewhere and this paradigm has been shown to produce pharmacologically relevant blood alcohol levels (e.g., Roberts et al. 1996; Roberts et al. 1999). Briefly, rats were restricted to three hours of water for three days and allowed access to the operant boxes where responding on the one extended lever resulted in the delivery of a saccharin solution. Thereafter, water restriction was discontinued. Ethanol concentrations increased from 5% to 8% to a final concentration of 10% over the following 20 days, with each concentration first being mixed with saccharin and then presented alone. During this saccharin fading procedure, both levers were extended, with one lever producing ethanol/saccharin and the other producing water. The levers associated with each solution were alternated between left and right on consecutive days. Rats were allowed to respond for 10% ethanol versus water for 4–6 weeks (until responding across three consecutive days varied less than 25% and preference for ethanol over water was at least 60%) before exposure to the vapor chambers.
Operant sessions were 30 minutes during the training phase and were conducted five days per week between 9:00 AM and 12:00 PM. Extended operant self-administration test sessions (12 hr) were conducted in Experiments 1 and 2. These were initiated at 6:00 PM, the beginning of the rats’ active phase (dark cycle). Food was placed in each operant box and 10% ethanol and water were available contingent on lever pressing. Test sessions in Experiments 3 and 4 were conducted in a manner similar to those used during training (30 min).
Ethanol Vapor Chamber Procedure
Ethanol vapor exposure is a reliable technique for inducing ethanol dependence in that blood alcohol levels can be tightly controlled and animals are free-moving and gain weight normally (Rogers et al. 1979). The ethanol vapor chambers and procedure were similar to those described by Rogers and colleagues (1979). Two standard rat cages were housed in each separate, sealed clear plastic chamber into which ethanol vapor, or air for control chambers, was independently introduced. Ethanol vapor was created by dripping 95% ethanol into 2000 ml Erlenmeyer vacuum flasks kept at 50°C on a warming tray. Air was blown over the bottom of the flask at a rate of 11 L/min to vaporize the ethanol. Concentrations of ethanol vapor were adjusted by varying the rate at which ethanol was pumped into the flask and ranged from 22 mg/L to 27 mg/L.
Target blood alcohol levels were 150–200 mg% across the 2–4 week exposure time. This paradigm has been shown to produce physical dependence as evidenced by the appearance of observable withdrawal symptoms upon removal from the chambers (Roberts et al. 1996). Blood was sampled for blood alcohol level determination every three days during vapor exposure and upon removal from the chambers. Tail blood (0.5 ml) was collected into heparinized Eppendorf tubes. After centrifugation, the plasma was extracted with trichloroacetic acid and assayed for ethanol content using the NAD-ADH enzyme spectrophotometric method (Sigma Chemical Co., St. Louis, MO).
Experiment 1
The purpose of this experiment was to determine whether increased responding for ethanol persisted beyond the acute withdrawal phase. It was shown previously that rats trained to lever press for ethanol and then exposed to ethanol vapor for two weeks, self-administered more ethanol in operant boxes than control rats exposed to air across an entire 12-hr withdrawal test session (Roberts et al. 1996). The difference between ethanol- and air-exposed rats became more striking with repeated tests, presumably as the association between ethanol drinking and relief of withdrawal symptoms strengthened. In this experiment, rats were tested in 12-hr sessions two, four, and eight weeks following removal from the vapor chambers, in addition to testing twice during withdrawal. It was hypothesized that previously ethanol vapor-exposed rats would show long-lasting increases in ethanol self-administration due to an alteration in hedonic set point.
Eight rats were trained to lever press for 10% ethanol and exposed to ethanol vapor (n = 4) or control air (n = 4). Groups were matched based on the final three days of 30-min operant testing. Following two weeks of vapor exposure, all rats were removed and placed immediately in operant boxes for a 12-hr overnight (active phase of circadian cycle) ethanol self-administration session. The rats were then re-exposed to the vapor chambers for five days and retested in a second 12-hr overnight session. Following this test, the rats were left out of the vapor chambers and not disturbed, except for routine husbandry, for two weeks. At this time, another 12-hr operant ethanol self-administration session was conducted. The rats were retested in this manner at 4 and 8 weeks following removal from the vapor chambers with no exposure to the operant boxes between these retest sessions (see Table 1) .
Experiment 2
The purpose of Experiment 2 was to determine whether prior exposure to ethanol vapor would lead to increased ethanol self-administration even if the rats were not tested previously during early withdrawal. In this case, rats would be tested in the absence of potential learned associations between lever pressing for ethanol and relief from early withdrawal symptoms. Again, the hypothesis was that rats previously exposed to ethanol vapor should have greater motivation to lever press for ethanol following protracted withdrawal.
Twenty eight rats were trained to lever press for 10% ethanol. On the final day of training, a 12-hr overnight test in the operant boxes was conducted in order to control for the extra test given to the rats during withdrawal in Experiment 1, as well as to select groups showing indistinguishable patterns of responding. The rats were then exposed for two weeks to ethanol vapor (n = 13) or control air (n = 15). One rat in the ethanol vapor group was lost due to over-intoxication (BAL> 300 mg%). Following removal from the vapor chambers, the rats were not disturbed for two weeks at which time they were tested in a second 12-hr overnight session in the operant self-administration boxes. This was repeated four and eight weeks following removal from the vapor chambers (see Table 1).
Experiment 3
The purpose of this experiment was to examine the effect of prior exposure to chronic ethanol vapor and subsequent abstinence on the resumption of daily 30-min test sessions. The results of Experiment 2 suggested that the only difference between the groups following protracted abstinence (when rats were not tested during early withdrawal) was in the first hour of retests. Therefore, the purpose of this experiment was to examine the effects of chronic ethanol exposure and a two week period of abstinence on the alcohol deprivation effect and its recovery to stable baseline responding. In addition, these rats were scored for ethanol withdrawal severity in order to confirm that the ethanol vapor chamber protocol was producing physical dependence as had been found previously (Roberts et al. 1996).
Twelve rats were trained to lever press for 10% ethanol and then half were exposed to ethanol vapor (n = 6) and half were exposed to air (n = 6) for two weeks. Eight hours following removal from the chambers, the rats were tested for ethanol withdrawal symptoms. Rats were scored from 0 (undetectable) to 2 (severe) in a test of ventromedial distal limb flexion and observed for tail stiffness and abnormal body posture. The sum of these three measures represented overall withdrawal severity, yielding a range from 0 to 6. More detail regarding this procedure has been previously published (Macey et al. 1996). Following this simple test, the rats were left undisturbed for two weeks and then daily 30-min operant ethanol self-administration sessions were resumed. Rats were tested for 10 consecutive days (see Table 1).
Experiment 4
The results of the previous experiments showed that there appears to be an enhancement of the alcohol deprivation effect and an increase to a new higher baseline of responding for ethanol in rats previously exposed to ethanol vapor, relatively to air exposed rats. Thus, the purpose of Experiment 4 was to further investigate this finding by allowing rats three cycles of abstinence and retesting following a single ethanol vapor exposure. In this experiment, rats were exposed to ethanol vapor or air for one month. Again, it was hypothesized that rats previously exposed to ethanol vapor would show enhanced alcohol deprivation effects and higher baseline levels of responding over several repeated tests.
Twelve rats were trained to lever press for 10% ethanol and then half were exposed to ethanol vapor (n = 6), whereas the other half was exposed to air (n = 6) for four weeks. Following removal from the chambers, the rats were left undisturbed for two weeks and then daily 30-min operant ethanol self-administration sessions were resumed for seven days. Rats were deprived of ethanol (not exposed to operant self-administration boxes) for three 1-week intervals separated by seven daily 30-min operant sessions in which 10% ethanol and water were available (see Table 1).
Data Analysis
Operant responding was analyzed by mixed factor 2-way analysis of variance. The between-subject factor was group (ethanol vapor versus air control) and the within-subject factor was time (hours in Experiments 1 and 2 and repeated daily sessions in Experiments 3 and 4). Each cycle of retest was analyzed separately. Interactions between group and either hour or session were investigated using simple main effects analyses.
RESULTS
Operant responding for ethanol by rats in the present experiments was similar to that found in previous experiments in which blood alcohol levels were determined. For example, at the completion of a 12-hr early withdrawal test session, similar to that used in Experiment 1, rats were found to have blood alcohol levels of approximately 100 mg% (Roberts et al. 1996). Baseline responding for ethanol in 30-min test sessions has been associated with blood alcohol levels of 25–30 mg%, whereas higher levels of responding similar to those observed presently following protracted withdrawal have been associated with blood alcohol levels of 40–80 mg% (Roberts et al. 1999).
Experiment 1
Baseline responding for ethanol was 21.1 ± 2.4 responses in the final 30-min training session prior to ethanol vapor exposure. Blood alcohol levels achieved by rats in the ethanol vapor chambers averaged 157 ± 19 mg% across the two weeks of exposure (time course data not shown). Body weights upon removal from the vapor chambers were 454 ± 51 g in the ethanol vapor group and 436 ± 21 g in the air control group.
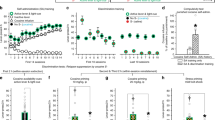
The results of this experiment are shown in Figure 1. The upper panels represent data from rats placed into the operant ethanol self-administration boxes immediately following removal from the vapor chambers for a first (left) and second (right) 12-hr test session. Rats exposed to ethanol vapor responded for ethanol significantly more than control rats in both the first (F[1.6] = 21.4, p < .01) and second (F[1,6] = 66.9, p < .001) sessions. The remaining panels of Figure 1 show similar tests in the same animals following progressively longer periods of abstinence. Ethanol vapor rats continued to respond more for ethanol than control rats at two weeks (F[1,6] = 9.3, p < .05), four weeks (F[1,6] = 15.1, p < .01), and eight weeks (F[1,6] = 7.4, p < .05) following removal from the vapor chambers. Asterisks indicate significant group differences as determined from simple effects analyses of the interactions between group and time for each test session. The insets to each panel, showing total ethanol consumption (12 hr) relative to body weights, depict the overall group differences in ethanol self-administration. In each case, rats exposed to the ethanol vapor chambers (either during early withdrawal or following protracted abstinence) responded for significantly more ethanol in g/kg over the entire 12-hr test session (p < .05).

Operant responding for oral ethanol across 12-hr test sessions in rats exposed to two weeks of ethanol vapor (n = 4) or air (n = 4). Rats were tested immediately upon removal from the ethanol vapor chambers and then again at 2, 4, and 8 weeks following removal. The insets to each panel depict total ethanol consumption relative to body weight for ethanol vapor-exposed (black bar) and air-exposed (white bar) rats. The numbers of ethanol deliveries and g/kg consumptions are represented as means ± SEM. The symbol * indicates a significant difference between the ethanol and control groups (p < .05)
These results suggest that the enhancement in operant responding for ethanol by rats exposed to chronic ethanol vapor which accompanies acute withdrawal is maintained over several weeks of protracted abstinence.
Experiment 2
Baseline responding for ethanol was 27.2 ± 3.8 responses in the final 30-min training session prior to ethanol vapor exposure. Blood alcohol levels of ethanol vapor-exposed rats averaged 149 ± 15 mg%. Body weights upon removal from the vapor chambers were 483 ± 20 g in the ethanol vapor group and 486 ± 18 g in the air control group.
The results of Experiment 2 are shown in Figure 2. In this experiment, rats were exposed to their first 12-hr operant ethanol self-administration test session prior to ethanol vapor or air exposure. These data were used to assign matched groups; therefore, as shown in the upper left panel of Figure 2, there were no group differences prior to vapor exposure. The remaining panels show similar tests in the same animals at various time points following removal from chronic vapor exposure. In contrast to the results of Experiment 1, there were no statistically significant main effects of group at any retest session, as is depicted by the overall ethanol consumption (12 hr) relative to body weights shown in the insets to each panel. There were, however, significant interactions between group and time at two weeks (F[11,275] = 3.1, p < .001) and four weeks (F[11,275] = 2.6, p < .01). Simple main effects analyses of these interactions revealed highly significant group differences in the first hour of both the 2-week (F[1,214] = 23.5, p < .0001) and 4-week (F[1,214] = 16.1, p < .0001) test. There were no differences involving group in the 8-week test. As depicted in the figure insets, there were no group differences in total ethanol intake in g/kg during any test session. The results of this experiment suggest that chronic ethanol exposure is associated with increased responding for ethanol even if rats are not tested during withdrawal. However, this effect was not sustained over the entire 12-hr period, but was only detected in the first hour of the test session.

Operant responding for oral ethanol across 12-hr test sessions in rats exposed to two weeks of ethanol vapor (n = 12) or air (n = 15). Rats were not tested immediately following removal from the vapor chambers, but were tested prior to vapor exposure both to equalize the testing between Experiments 1 and 2, as well as to select indistinguishable ethanol vapor and air groups. Rats were tested at 2, 4, and 8 weeks following removal from the vapor chambers. The insets to each panel depict total ethanol consumption relative to body weight for ethanol vapor-exposed (black bar) and air-exposed (white bar) rats. The numbers of ethanol deliveries and g/kg consumptions are represented as means ± SEM. The symbol * indicates a significant difference between the ethanol and control groups (p < .05)
Experiment 3
Baseline responding for ethanol was 26.3 ± 3.7 responses in the final 30-min training session prior to ethanol vapor exposure. Blood alcohol levels averaged 150 ± 6 mg% across the two weeks of ethanol vapor exposure. Body weights upon removal from the vapor chambers were 492 ± 21 g in the ethanol vapor group and 481 ± 16 g in the air control group. Withdrawal scores in ethanol vapor-treated rats averaged 3.9 ± 0.4 and in air controls were 0.57 ± 0.2. The scores of the ethanol vapor group are consistent with previous results (Roberts et al. 1996) and represent mild-to-moderate withdrawal severity.
The results of Experiment 3 are shown in Figures 3 and 4. Figure 3 shows operant responding for ethanol (top) and water (bottom) in daily 30-min sessions before vapor exposure and two weeks following removal from vapor chambers. Pre-vapor data were used to assign matched groups. Following abstinence, both groups showed an increase in ethanol self-administration consistent with the alcohol deprivation effect (p < .05). Importantly, ethanol exposed rats responded for significantly more ethanol than air controls following vapor exposure (F[1,12] = 13.5, p < .01). This group difference was significant on days 1, 2, 4, 5, and 6 (p < .05). There was no group difference in responding for water (F[1,12] = 0.01, p = .9).

Operant responding for oral ethanol (top) and water (bottom) across 10 days of 30-min test sessions in rats exposed to two weeks of ethanol vapor (n = 6) or air (n = 6). The three pre-vapor test sessions were used for group selection. Daily operant test sessions were resumed two weeks following removal from the vapor chambers. Numbers of deliveries are represented as means ± SEM. The symbol * indicates a significant difference between the ethanol and control groups (p < .05)

Cumulative operant responding for ethanol in the last baseline day (top), first retest day following two weeks of vapor exposure and two weeks of abstinence (middle), and tenth retest day (bottom) in ethanol vapor (n = 6) and air control (n = 6) rats. Each point represents the cumulative responding (mean ± SEM) in 5-min intervals. Ethanol intakes in g/kg are listed next to the right of the lines. The symbol * indicates a significant difference between the ethanol and control groups (p < .05)
Figure 4 shows cumulative responding for ethanol in the first (middle) and tenth (bottom) sessions following vapor exposure and abstinence. The last baseline session prior to vapor exposure is shown (top) for comparative purposes. On the first day post vapor exposure, air control rats showed a steeper cumulative response profile than during the baseline session, characteristic of the alcohol deprivation effect described in nondependent rats (Heyser et al. 1997). Ethanol vapor rats show an even more pronounced alcohol deprivation effect in the first retest session as indicated by a significant main effect of group (F[1,12] = 11.7, p < .01) and a significant group by time epoch interaction (F[5,60] = 3.8, p < .01). Simple main effects analysis of Day 1 data revealed significantly higher responding by ethanol vapor rats at all but the first time epoch (F[1,23] = 4.9–16.7, p < .05). On the tenth day post vapor exposure, there was no significant main effect of group, but the interaction between group and time epoch was statistically reliable (F[5,60] = 4.9, p < .001). Simple main effects analyses of Day 10 data revealed higher responding by ethanol vapor rats in the final two 5-min time intervals (F[1,20] = 4.9–5.8, p < .05). These results suggest that prior exposure to chronic ethanol vapor is associated with enhanced responding for ethanol across 10 daily test sessions.
Experiment 4
Baseline responding for ethanol was 19.2 ± 3.1 responses in the final 30-min training session prior to ethanol vapor exposure. Blood alcohol levels of ethanol vapor rats averaged 143 ± 15 mg% for the one-month exposure time. Body weights upon removal from the vapor chambers were 506 ± 17 g in the ethanol vapor group and 522 ± 16 g in the air control group.
The results of this experiment are shown in Figure 5. As in Experiment 3, the final three pre-vapor days are shown, followed by seven daily sessions subsequent to vapor exposure plus two weeks of abstinence. Three additional 7-day cycles of responding following one-week abstinence periods also are shown. Rats with a history of ethanol vapor exposure responded for significantly more ethanol than control rats across all four retest blocks (F[1,10] = 6.2, p < .05). There was an overall significant interaction between group and test session (F[6,60] = 2.9, p < .05), with simple main effects analyses revealing significant effects of group in the first three test sessions of the blocks (F[1,18] = 8.6–9.1, p < .01). The results of Experiment 4 are in accordance with those of the previous three experiments in that prior exposure to chronic ethanol vapor is associated with an increase in operant ethanol self-administration. As in Experiment 2, when rats were not tested during withdrawal, this effect appeared to be statistically reliable through the 4th or 5th week following removal from the vapor chambers.

Operant responding for oral ethanol across four cycles of seven days of 30-min test sessions separated by one week of ethanol deprivation. The three pre-vapor test sessions were used for group selection. The cycles of daily operant test sessions were initiated two weeks following removal from the vapor chambers. Numbers of deliveries are represented as means ± SEM. Ethanol intakes prior to vapor exposure were 0.39 ± 0.1 g/kg for the control group and 0.36 ± 0.1 for the ethanol vapor group. On the first day of each deprivation cycle intakes were 0.50–0.66 g/kg for control rats and 0.81–1. 2 g/kg for ethanol vapor exposed rats. On the last day of the experiment, the control rats responded for 0.48 ± 0.15 g/kg ethanol and the ethanol vapor exposed rats responded for 0.71 g/kg ethanol. The symbol * indicates a significant difference between the ethanol and control groups (p < .05)
DISCUSSION
The principal result of these experiments was that rats with a history of chronic ethanol exposure, sufficient to produce signs of physical dependence, showed persistent increases in operant ethanol self-administration during withdrawal and protracted abstinence. This is an important finding as it suggests that increases in ethanol self-administration can persist for 4–8 weeks following a single chronic ethanol exposure sufficient to produce dependence. The increase was anywhere from 30% to 100% above baseline, and, based on previous data would result in an increase in blood alcohol levels from about 25–30 mg% to 40–80 mg% in 30-min sessions (Roberts et al. 1999). In the first experiment, rats were allowed access to the operant boxes during two early withdrawal experiences and then retested following more protracted periods of abstinence. In Experiment 2, rats were tested prior to vapor exposure to examine baseline response rates across a 12-hr session and to select equivalent groups and then not tested again until following protracted periods of abstinence. These experiments were carried out using extended 12-hr operant test sessions. Experiments 3 and 4 were designed to examine operant responding for ethanol across daily 30-min sessions following periods of protracted abstinence in order to characterize the persistence of the changes observed in Experiments 1 and 2 and more closely examine the alcohol deprivation effect.
The differences between the findings of Experiments 1 and 2 are quite intriguing and suggest that experience with ethanol self-administration during the early withdrawal period leads to a more robust increase in ethanol self-administration under conditions of protracted abstinence. This may be due to the development of an association between ethanol drinking and alleviation of withdrawal signs which enhances the reinforcing efficacy of ethanol. If animals are allowed to self-administer ethanol during repeated withdrawals, they show more robust increases in preference for ethanol (Hunter et al. 1974; Roberts et al. 1996). This has important implications for relapse in human alcoholics as it suggests that violations of abstinence during the early withdrawal phase may increase the magnitude of subsequent lapses. Certainly, in human studies there is strong evidence that the longer a recovering alcoholic can stay abstinent, the lower the chance for relapse (De Soto et al. 1989). This may primarily be due to the recovery from the protracted withdrawal syndrome and repair of social and occupational functions, but it also highlights the importance of coping strategies as a mechanism to relearn alternate behaviors to alleviate negative affect and to extinguish alcohol drinking as the primary mechanism to this end. It should be noted, however, that the rats used in Experiment 1 also had an additional withdrawal experience and five more days of vapor exposure as well as slightly lower pre-vapor levels of responding. These factors also may have contributed to the increased protracted abstinence self-administration observed in Experiment 1.
In addition to the development of learned associations between ethanol drinking and withdrawal relief, more pronounced withdrawal symptomatology with repeated withdrawal episodes also may contribute to increases in the reinforcing effects of ethanol. It is well established that the amount and duration of ethanol exposure prior to abstinence is an important determinant of withdrawal severity, and there is striking evidence in the literature that the experience of repeated withdrawal is associated with enhancements in withdrawal symptomatology (Brown et al. 1988; Lechtenberg and Worner 1991; Booth and Blow 1993; Becker 1999). For example, in a large national study of alcoholic men it was found that the occurrence of seizures during detoxification was associated with significantly greater numbers of prior withdrawals (Booth and Blow 1993). Such enhancements in seizure susceptibility with repeated withdrawal episodes has been modeled in animals (Branchey et al. 1971; Baker and Cannon 1979; Becker and Hale 1993; Becker 1994). Interestingly, an enhancement in anxiety-like behavior as determined by elevated plus maze performance was found following repeated periods of deprivation from ethanol in rats not showing classical ethanol withdrawal symptomatology (Hölter et al. 1998). Thus, it is likely that repeated bouts of ethanol exposure and abstinence may result in more permanent changes in the reinforcing effects of ethanol than those found herein. For example, responding for ethanol immediately following removal from alcohol vapor chambers became more consistent over repeated withdrawals (Roberts et al. 1996).
The results replicate the finding of increased ethanol self-administration during early withdrawal shown previously (Roberts et al. 1996) and also clearly demonstrate the presence of the alcohol deprivation effect in both control and ethanol vapor exposed rats. Experiments 3 and 4 were designed to more closely examine the alcohol deprivation effect in previously dependent animals. Based on a hypothesis of an allostatic change in reward set point, it would be predicted that both deprivation levels of ethanol self-administration, as well as baseline levels would be elevated. Control rats showed a typical transient increase in ethanol self-administration in the first session following a period of abstinence, however this deprivation effect was larger in rats having a history of ethanol dependence. Thus, the alcohol deprivation effect does not appear to be changed in any qualitative manner following dependence, but rather an overall increase in ethanol self-administration has occurred. The level of ethanol self-administration observed during protracted abstinence in this population of genetically heterogeneous rats is similar to that observed under similar conditions, but with no deprivation periods, in rats genetically selected for preference for alcohol (Li and McBride 1995; Files et al. 1998; Samson et al. 1998).
The persistent increase in operant responding for ethanol following protracted abstinence is consistent with an elevation in the ethanol reward set point or threshold for ethanol reward of the animals. A hypothesis of an allostatic set point shift predicts no change in sensitivity to ethanol per se, but rather that a change in the reward state even in the absence of drug has occurred (Koob and Le Moal 1997). This model of allostasis could explain the enhancement of vulnerability to relapse following the development of dependence, as reward function in the drug-free state is hypothesized to be decreased below preaddicted levels. Thus, the motivation to consume ethanol in order to even achieve euthymia or normal reward function would be quite great. Alternatively, the increase in ethanol responding in rats with a history of chronic ethanol exposure may be due to the development of tolerance to some effect(s) of ethanol. In this framework, more ethanol is taken by the animals in order to achieve the same effect because a decrease in sensitivity to ethanol has occurred (Kalant et al. 1971). It is not likely that tolerance can explain the present results, however, because rats receiving the same training and ethanol vapor histories showed very different response patterns depending on whether they were tested during early withdrawal or not (Figure 1 vs. Figure 2). In addition, the increased responding observed in the first hour of the extended sessions or the first session of the repeated daily tests, while declining within each session or set of sessions, was regenerated following periods of abstinence.
These findings are not compatible with a change in sensitivity to ethanol, but rather are more convincingly explained by changes in motivational set-point which would be greatest following periods of abstinence and also enhanced by the association between ethanol self-administration and the alleviation of withdrawal symptoms. Both allostasis and tolerance are descriptive constructs which have been used to explain the changes that occur in drug and alcohol self-administration with chronic use, and it is certainly possible that they are not mutually exclusive, but rather both play a role in the effects observed herein (for a more detailed discussion of this point see Ahmed and Koob 1999).
The present results also have potentially important implications for understanding the characteristics and underlying neurobiological mechanisms of relapse. A long-lasting change in ethanol self-administration has been demonstrated, making it possible to examine the relapse phenomenon after ethanol has been removed for varying periods. For example, the putative role of anxiety in ethanol drinking following protracted abstinence can be more directly examined. In addition, potential pharmacotherapies can be tested in this preclinical model, as the course of protracted abstinence (i.e., the detoxification period) is an obvious target for anti-relapse interventions. There is substantial evidence that chronic ethanol exposure results in changes in a variety of neuronal systems such as those involving adenylate cyclase, gamma-aminobutyric acid (GABA), norepinephrine, serotonin, vasopressin, glutamate (Tabakoff and Hoffman 1992) as well as dopamine and corticotropin-releasing factor (CRF) (Nutt and Glue 1990).
To date, there is evidence that changes in dopamine, GABA, endogenous opioids, glutamate and CRF may be associated with the transition from controlled alcohol use to alcohol dependence (Roberts and Koob 1997; Koob et al. 1998). For example, CRF is increased in the amygdala during early ethanol withdrawal in rats (Merlo-Pich et al. 1995) and the administration of CRF receptor antagonists into this brain region reverses the anxiety-like behaviors measured during early ethanol withdrawal (Rassnick et al. 1993). The CRF system, which mediates many behavioral responses to stress, is of particular interest as it may be critical for the negative affect and anxiety-like effect associated with protracted abstinence. The models described herein will allow for the more direct characterization of the role of different neurochemical systems in ethanol self-administration following protracted abstinence and enhance our understanding of the factors, environmental, genetic, and neuropharmacological, that lead to relapse.
References
Ahlenius S, Engel J . (1974): Behavioral stimulation induced by ethanol withdrawal. Pharmacol Biochem Behav 2: 847–850
Ahmed SH, Koob GF . (1998): Transition from moderate to excessive drug intake: Change in hedonic set point. Science 282: 298–300
Ahmed SH, Koob GF . (1999): Long-lasting increase in the set point for cocaine self-administration after escalation in rats. Psychopharmacology 146: 303–312
U.S. Department of Health and Human Services. (1990): Seventh Special Report to the U.S. Congress on Alcohol and Health. Rockville, MD, U.S. Department of Health and Human Services.
Alling C, Balldin J, Bokström K, Gottfries CG, Karlsson I, Langström G . (1982): Studies on duration of a late recovery period after chronic abuse of ethanol: A cross-sectional study of biochemical and psychiatric indicators. Acta Psychiatry Scand 66: 384–397
Annis HM, Sklar SM, Moser AE . (1998): Gender in relation to relapse crisis situations, coping, and outcome among treated alcoholics. Addict Behav 23: 127–131
Baker TB, Cannon DS . (1979): Potentiation of ethanol withdrawal by prior dependence. Psychopharmacology 60: 105–110
Becker HC . (1994): Positive relationship between the number of prior ethanol withdrawal episodes and the severity of subsequent withdrawal seizures. Psychopharmacology 116: 26–32
Becker HC . (1999): Alcohol withdrawal: Neuroadaptation and sensitization. CNS Spectrums 4: 38–65
Becker HC, Hale RL . (1993): Repeated episodes of ethanol withdrawal potentiate the severity of subsequent withdrawal seizures: An animal model of alcohol withdrawal “kindling”. Alcohol Clin Exp Res 17: 94–98
Begleiter H . (1975): Ethanol consumption subsequent to physical dependence. Adv Exp Med Biol 59: 373–378
Begleiter H, Porjesz B . (1979): Persistence of a “subacute withdrawal syndrome” following chronic ethanol intake. Drug Alcohol Depend 4: 353–357
Booth BM, Blow FC . (1993): The kindling hypothesis: Further evidence from a U.S. national study of alcoholic men. Alcohol Alcohol 28: 593–598
Branchey M, Rauscher G, Kissin B . (1971): Modifications in the response to alcohol following the establishment of physical dependence. Psychopharmacology 22: 314–322
Brown G, Jackson A, Stephens DN . (1998): Effects of repeated withdrawal from chronic ethanol on oral self-administration of ethanol on a progressive ratio schedule. Behav Pharmacol 9: 149–161
Brown ME, Anton RF, Malcolm R, Ballenger JC . (1988): Alcohol detoxification and withdrawal seizures: Clinical support for a kindling hypothesis. Biol Psychiatry 23: 507–514
Cappell H, LeBlanc AE . (1981): Tolerance and physical dependence: Do they play a role in alcohol and drug self-administration? Res Adv Alcohol Drug Probl 6: 159–196
Denoble VJ, Begleiter H . (1978): Alcohol self-administration in monkeys (Macaca radiata): The effects of prior alcohol exposure. Pharmacol Biochem Behav 8: 391–397
De Soto CB, O'Donnell WE, Allred LJ, Lopes CE . (1985): Symptomology in alcoholics at various stages of abstinence. Alcohol Clin Exp Res 9: 505–512
De Soto CB, O'Donnell WE, De Soto JL . (1989): Long-term recovery in alcoholics. Alcohol Clin Exp Res 13: 693–697
Deutsch JA, Koopmans HS . (1973): Preference enhancement for alcohol by passive exposure. Science 179: 1242–1243
Deutsch JA, Walton NY . (1977): A rat alcoholism model in a free choice situation. Behav Biol 19: 349–360
Edwards G . (1990): Withdrawal symptoms and alcohol dependence: Fruitful mysteries. Br J Addict 85: 447–461
Files FJ, Samson HH, Denning CE, Marvin S . (1998): Comparison of alcohol-preferring and nonpreferring selectively bred rat lines. II. Operant self-administration in a continuous access situation. Alcohol Clin Exp Res 22: 2147–2158
Grant I, Reed R, Adams KM . (1987): Diagnosis of intermediate-duration and subacute organic mental disorders in abstinent alcoholics. J Clin Psychiatry 48: 319–323
Hershon HI . (1977): Alcohol withdrawal symptoms and drinking behavior. J Stud Alcohol 38: 953–971
Heyser CJ, Schulteis G, Koob GF . (1997): Increased ethanol self-administration after a period of imposed ethanol deprivation in rats trained in a limited access paradigm. Alcohol Clin Exp Res 21: 784–791
Hölter SM, Engelmann M, Kirschke C, Liebsch G, Landgraf R, Spanagel R . (1998): Long-term ethanol self-administration with repeated ethanol deprivation episodes changes ethanol drinking pattern and increases anxiety-related behavior during ethanol deprivation in rats. Behav Pharmacol 9: 41–48
Hunter BE, Walker DW, Riley JN . (1974): Dissociation between physical dependence and volitional ethanol consumption: Role of multiple withdrawal episodes. Pharmacol Biochem Behav 2: 523–529
Kalant H, LeBlanc AE, Gibbins RJ . (1971): Tolerance to and dependence on some nonopiate psychotropic drugs. Pharmacol Rev 23: 135
Kissin B . (1979): Biological investigations in alcohol research. J Stud Alcohol(Suppl) 8: 146–181
Koob GF, Le Moal M . (1997): Drug abuse: Hedonic homeostatic dysregulation. Science 278: 52–58
Koob GF, Roberts AJ, Schulteis G, Parsons LH, Heyser CJ, Hyytiä P, Merlo-Pich E, Weiss F . (1998): Neurocircuitry targets in ethanol reward and dependence. Alcohol Clin Exp Res 22: 3–9
Lechtenberg R, Worner TM . (1991): Relative kindling effect of detoxification and non-detoxification admissions in alcoholics. Alcohol Alcohol 26: 221–225
Li T-K, McBride WJ . (1995): Pharmacogenetic models of alcoholism. Clin Neurosci 3: 182–188
Macey DJ, Schulteis G, Heinrichs SC, Koob GF . (1996): Time-dependent quantifiable withdrawal from ethanol in the rat: Effect of method of dependence induction. Alcohol 13: 163–170
Meisch RA . (1983): Relationship between physical dependence on ethanol and reinforcing properties of ethanol in animals. In Cicero TJ (ed), Ethanol Tolerance and Dependence: Endocrinological Aspects, [NIAAA Research Monograph vol. 13]. Rockville, MD, National Institute on Alcohol Abuse and Alcoholism, pp 27–32
Meisch RA . (1994): Alcohol self-administration by experimental animals. Res Adv Alcohol Drug Probl 8: 23–45
Merlo-Pich E, Lorang M, Yeganeh M, Rodriguez de Fonseca F, Raber J, Koob GF, Weiss F . (1995): Increase of extracellular corticotropin-releasing factor-like immunoreactivity levels in the amygdala of awake rats during restraint stress and ethanol withdrawal as measured by microdialysis. J Neurosci 15: 5439–5447
Myers RD, Stoltman WP, Martin GE . (1972): Effects of ethanol dependence induced artificially in the Rhesus monkey on the subsequent preference for ethyl alcohol. Physiol Behav 9: 43–48
Nutt DJ, Glue P . (1990): Neuropharmacological and clinical aspects of alcohol withdrawal. Ann Med 22: 275–281
Rassnick S, Heinrichs SC, Britton KT, Koob GF . (1993): Microinjections of a corticotropin-releasing factor antagonist into the central nucleus of the amygdala reverses anxiogenic-like effects of ethanol withdrawal. Brain Res 605: 25–32
Roberts AJ, Cole M, Koob GF . (1996): Intra-amygdala muscimol decreases operant ethanol self-administration in dependent rats. Alcohol Clin Exp Res 20: 1289–1298
Roberts AJ, Koob GF . (1997): The neurobiology of addiction. Alcohol Health Res World 21: 101–106
Roberts AJ, Heyser CJ, Koob GF . (1999): Operant self-administration of sweetened versus unsweetened ethanol: Effects on blood alcohol levels. Alcohol Clin Exp Res 23: 1151–1157
Roelofs SMGJ . (1985): Hyperventilation, anxiety, craving for alcohol: a subacute alcohol withdrawal syndrome. Alcohol 2: 501–505
Rogers J, Wiener SG, Bloom FE . (1979): Long-term ethanol administration methods for rats: Advantages of inhalation over intubation of liquid diets. Behav Neural Biol 27: 466–486
Samson HH . (1986): Initiation of ethanol reinforcement using a sucrose-substitution procedure in food- and water-sated rats. Alcohol Clin Exp Res 10: 436–442
Samson HH, Falk JL . (1974): Alteration of fluid preference in ethanol-dependent animals. J Pharmacol Exp Ther 190: 365–376
Samson HH, File FJ, Denning C, Marvin S . (1998): Comparison of alcohol-preferring and nonpreferring selectively bred rat lines. I. Ethanol initiation and limited access operant self-administration. Alcohol Clin Exp Res 22: 2133–2146
Schulteis G, Hyytïa P, Heinrichs SC, Koob GF . (1996): Effects of chronic ethanol exposure on oral self-administration of ethanol or saccharin by Wistar rats. Alcohol Clin Exp Res 20: 164–171
Sinclair JD, Senter RJ . (1967): Increased preference for ethanol in rats following alcohol deprivation. Psychon Sci 8: 11–12
Spanagel R, Hölter SM, Allingham K, Landgraf R, Zieglgänsberger W . (1996): Acamprosate and alcohol. I. Effects on alcohol intake following alcohol deprivation in the rat. Eur J Pharmacol 305: 39–44
Sterling P, Eyer J . (1988): Allostasis: A new paradigm to explain arousal pathology. In Fisher S, Reason J (eds), Handbook of Life Stress, Cognition and Health, Chichester, England, John Wiley & Sons, Ltd, pp 629–649
Tabakoff B, Hoffman PL . (1992): Alcohol: Neurobiology. In Lowinson JH, Ruiz P, Millman RB (eds), Substance Abuse: A Comprehensive Textbook. Baltimore, Williams and Wilkins, pp 152–185
Winger G . (1988): Effects of ethanol withdrawal on ethanol-reinforced responding in Rhesus monkeys. Drug Alcohol Dep 22: 235–240
Acknowledgements
The authors wish to thank Tess Kimber for her technical expertise, Dr. Serge Ahmed for fruitful discussions, and Mike Arends for his expert editorial assistance. These studies were supported by National Institute of Health grants from the National Institute on Alcohol Abuse and Alcoholism (AA05457, AA06420, and AA08459). This manuscript is publication 12165-NP of The Scripps Research Institute.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Roberts, A., Heyser, C., Cole, M. et al. Excessive Ethanol Drinking Following a History of Dependence: Animal Model of Allostasis. Neuropsychopharmacol 22, 581–594 (2000). https://doi.org/10.1016/S0893-133X(99)00167-0
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1016/S0893-133X(99)00167-0
Keywords
This article is cited by
-
Hypodopaminergic state of the nigrostriatal pathway drives compulsive alcohol use
Molecular Psychiatry (2023)
-
Structured tracking of alcohol reinforcement (STAR) for basic and translational alcohol research
Molecular Psychiatry (2023)
-
Translational opportunities in animal and human models to study alcohol use disorder
Translational Psychiatry (2021)
-
Vulnerability factors for mephedrone-induced conditioned place preference in rats—the impact of sex differences, social-conditioning and stress
Psychopharmacology (2021)
-
Anti-relapse neurons in the infralimbic cortex of rats drive relapse-suppression by drug omission cues
Nature Communications (2019)
