
Abstract
It has been hypothesized that alcohol addiction is mediated, at least in part, by specific γ-aminobutyric acidA (GABAA) receptors within the ventral pallidum (VP). Among the potential GABAA receptor isoforms regulating alcohol-seeking behaviors within the VP, the GABAA α1 receptor subtype (GABAA1) appears pre-eminent. In the present study, we developed β-carboline-3-carboxylate-t-butyl ester (βCCt), a mixed agonist–antagonist benzodiazepine (BDZ) site ligand, with binding selectivity at the A1 receptor to explore the functional role of VPA1 receptors in the euphoric properties of alcohol. The in vivo actions of βCCt were then determined following microinfusion into the VP, a novel alcohol reward substrate that primarily expresses the A1 receptor. In two selectively bred rodent models of chronic alcohol drinking (HAD-1, P rats), bilateral microinfusion of βCCt (0.5–40 μg) produced marked reductions in alcohol-reinforced behaviors. Further, VP infusions of βCCt exhibited both neuroanatomical and reinforcer specificity. Thus, no effects on alcohol-reinforced behaviors were observed following infusion in the nucleus accumbens (NACC)/caudate putamen (CPu), or on response maintained by saccharin. Parenteral-administered βCCt (1–40 mg/kg) was equally effective and selective in reducing alcohol-reinforced behaviors in P and HAD-1 rats. Additional tests of locomotor activity revealed that βCCt reversed the locomotor sedation produced by both chlordiazepoxide (10 mg/kg) and EtOH (1.25 g/kg), but was devoid of intrinsic effects when given alone. Studies in recombinant receptors expressed in Xenopus oocytes revealed that βCCt acted as a low-efficacy partial agonist at α3β3γ2 and α4β3γ2 receptors and as a low-efficacy inverse agonist at α1β3γ2, α2β3γ2, and α5β3γ2 receptors. The present study indicates that βCCt is capable of antagonizing the reinforcing and the sedative properties of alcohol. These anti-alcohol properties of βCCt are primarily mediated via the GABAA1 receptor. βCCt may represent a prototype of a pharmacotherapeutic agent to effectively reduce alcohol drinking behavior in human alcoholics.
Similar content being viewed by others


INTRODUCTION
Alcoholism (ie alcohol addiction and dependence) can be defined, in part, as a maladaptive pattern of alcohol abuse, leading to clinically significant impairment (DSM-IV, 2000). There is a need for markedly increased amounts of alcohol to achieve intoxication. Further, because a great deal of time is spent by the alcoholic in activities trying to obtain alcohol, in many cases one's family structure and occupational stability is adversely affected (NIAAA, 1997; Kessler et al, 1997). Substantial progress has been made with rodent models in emulating the clinical criteria for alcoholism (Cloninger, 1987; McBride and Li, 1998). With the exception of psychosocial and cultural factors that influence drinking, rodent models, in particular, the alcohol-preferring (P) rats have been suggested to satisfy the DSM-IV criteria for alcoholism ‘reasonably well’ (for a detailed description, see McBride and Li, 1998).
While advances have been made in the development of novel therapies to treat alcoholism (O'Malley et al, 1992; Volpicelli et al, 1992; Kranzler, 2000; Spanagel and Zieglgansberger, 1997), alcohol-dependent individuals represent a heterogeneous group (Cloninger, 1987; Li et al, 1991; Li, 2000), and it is unlikely that a single pharmacological treatment will be effective for all alcoholics. Hence, a better understanding of the neuromechanisms that regulate alcohol-seeking behaviors and the design of clinically safe and effective drugs that reduce alcohol addiction and dependence remain high priorities (Kranzler, 2000; Johnson and Ait-Daoud, 2000). While the precise neuromechanisms regulating alcohol-seeking behaviors remain unknown, there is now compelling evidence that the γ-aminobutyric acidA (GABAA) receptors within the striatopallidal and extended amygdala system are involved in the ‘acute’ reinforcing actions of alcohol (Koob, 1998; Koob et al, 1998; June et al, 1998c; McBride and Li, 1998). The striatopallidal and extended amygdala system include the sublenticular extended amygdala (substantia innominata-ventral pallidum (VP)), the shell of the nucleus accumbens (NACC), and the central nucleus of the amygdala (Heimer et al, 1991; Heimer and Alheid, 1991). Among the potential GABAA receptor isoforms within the VP capable of regulating alcohol-seeking behaviors, the GABAA receptors containing the α1 receptor subtype (GABAA1) appear to be the most logical candidate. Thus, Criswell et al (1993), (1995) observed that acute alcohol administration selectively enhanced the effects of ionotophoretically applied GABA in the VP. However, no effects were seen in the septum, VTA, and CA1 hippocampus. Further, a positive correlation was observed between alcohol-induced GABA enhancement and [3H]zolpidem binding (an A1 subtype selective agonist). Other investigators have identified a dense reciprocal projection from the VP to the NACC (Nauta et al, 1978; Zahm and Heimer, 1988; Groenewegen et al, 1993), and many of these have been found to be GABAergic neurons (Mogenson and Nielson, 1983; Kuo and Chang, 1992; Churchill and Kalivas, 1994). The NACC is well established as a substrate that regulates the reinforcing properties of abused drugs (Koob, 1998; Koob et al, 1998). Finally, immunohistochemical (Turner et al, 1993; Fritschy and Möhler, 1995) and in situ hybridization studies (Churchill et al, 1991; Wisden et al, 1992; Duncan et al, 1995) have demonstrated that the VP contains one of the highest concentrations of mRNA encoding the A1 subunit in the CNS. These findings, together with pharmacological studies suggesting that the VP plays a role in reward-mediated behaviors of psychostimulants and opiates (Hubner and Koob, 1990; Napier and Chrobak, 1992; Churchill and Kalivas, 1994; Gong et al, 1996, 1997), led us to hypothesize that the A1 containing GABAA receptors regulate alcohol-motivated behaviors.
To test this hypothesis, we developed β-carboline-3-carboxylate-t-butyl ester (βCCt), a mixed benzodiazepine (BDZ) agonist–antagonist with binding selectivity at the A1 receptor. Behavioral studies in several species (eg rats, mice, and primates) show that βCCt is a BDZ antagonist, exhibiting competitive binding site interaction with BDZ agonists over a broad range of doses (Shannon et al, 1984; Griebel et al, 1999; Cox et al, 1998; Carroll et al, 2001; Rowlett et al, 2001; Paronis et al, 2001). Other studies show that βCCt produces anxiolytic effects in rodents (Carroll et al, 2001) and potentiates the anticonflict response induced by A1 subtype ligands in primates (Paronis et al, 2001). Thus, βCCt displays an agonist or antagonist profile depending on the behavioral task, species, and dose employed. Studies of recombinant receptors show that βCCt exhibits a >10-fold selectivity for the GABAA1 over the A2 and A3 receptors, and a >110-fold selectivity for the A1 over the A5 subtype (Cox et al, 1995). Hence, βCCt exhibits the greatest binding selectivity of the currently available A1 receptor ligands (Sanger et al, 1994; McKernan et al, 2000; Cox et al, 1998).
In the present study, in vitro studies were conducted in recombinant GABAA1–A5 receptors in Xenopus oocytes to determine the efficacy of βCCt. Next, a series of in vivo studies were conducted to examine the effects of βCCt to reduce alcohol responding following parenteral injections and direct infusions into the VP. The degree of neuroanatomical specificity in modulating alcohol drinking was examined following both bilateral and unilateral control injections of βCCt into the NACC/caudate putamen (Cpu). The specificity of βCCt on alcohol-induced responding was evaluated by determining its effects in P rats, whose response rates for EtOH (10% v/v) and saccharin solutions (0.05% w/v) were similar at basal levels. The abilities of βCCt were also examined on a caloric sucrose reward. Finally, since the GABAA1 receptor isoform has recently been implicated in the sedative effects of BDZs (Rudolf et al, 1999; McKernan et al, 2000; Löw et al, 2000), we tested the hypothesis that the GABAA1 receptor plays a role in the sedation produced by an intoxicating dose of alcohol (1.25 g/kg). Chlordiazepoxide was used as a reference BDZ agonist. Using the nomenclature recently recommended by the International Union of Pharmacology (IUPHAR), α1–α6 containing GABAA receptors are referred to in the present study as GABAA1–A6 receptors (Barnard et al, 1998).
MATERIALS AND METHODS
Chemistry and Molecular Biology
Synthesis of βCCt
βCCt (see Figure 1), an A1 subtype ligand, was synthesized by modification of the prototypical β-carboline, βCCE, using a previously developed method for t-butyl ester synthesis (see Cox et al, 1995, 1998).

Synthesis and structure of βCCt.
Xenopus oocyte expression assay
Xenopus laevis frogs were purchased from Xenopus-1 (Dexter, MI). Collagenase B was obtained from Boehringer Mannheim (Indianapolis, IN), and GABA from RBI (Natick, MA). All compounds were prepared as a 10 -mM stock solution in EtOH and stored at –20°C.
cDNA clones: The rat GABAA receptor A1, A5, and γ2 subunit clones were gifts from H.Luddens (Department of Psychiatry, University of Mainz, Germany). The rat GABAA receptor β3 subunit clone was a gift from L Mahan (NINDS, NIH). Capped cRNA was synthesized from linearized template cDNA encoding the subunits using mMESSAGE mMACHINE kits (Ambion, Austin, TX). Oocytes were injected with the α, β, and γ subunits in a 1 : 1 : 1 molar ratio as determined by UV absorbance. Mature X. laevis frogs were anesthetized by submersion in 0.1% 3-aminobenzoic acid ethyl ester, and the oocytes were surgically removed. Follicle cells were removed by treatment with collagenase B for 2 h. Each oocyte was injected with 50–100 ng of cRNA in 50 nl of water and incubated at 19°C in modified Barth's saline (88 mM NaCl, 1 mM KCl, 2.4 mM NaHCO3, 0.41 mM CaCl2, 0.82 mM MgSO4, 100 μg/ml gentamicin, and 15 mM HEPES, pH 7.6). Oocytes were recorded from after 3 to 10 days postinjection. Oocytes were perfused at room temperature in a Warner Instruments oocyte recording chamber #RC-5/18 (Hamden, CT) with a perfusion solution (115 mM NaCl, 1.8 mM CaCl2, 2.5 mM KCl, 10 mM HEPES, pH 7.2). The perfusion solution was gravity fed continuously at a rate of 15 ml/min (see Harvey et al, 1997). Compounds were diluted in the perfusion solution, and applied until after a peak current was reached.
Current responses to GABA application were measured under a two-electrode voltage clamp, at a holding potential of −60 mV. Data were collected using a GeneClamp 500 amplifier and Axoscope software (Axon Instruments, Foster City, CA). GABA concentration–response curves for the GABAA receptor subunit combinations were constructed by normalizing responses to a low concentration of GABA to minimize variability, and then renormalized to the maximal response for comparison. Concentration–response data were fitted to a four-parameter logistic using GraphPad Prism, and the EC50 for each receptor subtype was determined. The peak whole cell current responses of a voltage-clamped oocyte to an EC50 concentration of GABA in the presence of saturating (1–10 μM) concentrations of modulators are reported as a percentage of the peak response to GABA alone (‘percent GABA response’ or ‘% control’) (for details, see June et al, 2001).
The efficacy of βCCt was measured by peak whole cell current responses in the Xenopus oocyte assay to an EC50 concentration of GABA at saturating (1–10 μM) concentrations of the ligand. The efficacies of nonselective, competitive BDZ antagonists (eg Ro15-1788, ZK 93426) were also evaluated.
Neurobehavioral Studies
Subjects
General: The alcohol-preferring (P) and high-alcohol drinking (replicate line #1) (HAD-1) rats were used to model the human condition of alcohol abuse; both rat lines are accepted as animal models of the acute reinforcing effects of alcohol in humans to the satisfaction of the alcohol research community (Cloninger, 1987; Lumeng et al, 1995; McBride and Li, 1998). Rats were obtained from the Alcohol Research Center at Indiana University School of Medicine. Female P and HAD-1 rats were used in all experiments; however, due to the large number of P (n=128) and HAD-1 (n=98) rats within the experimental design (total N=226), it was virtually impossible to obtain all P or HAD-1 rats of a single generation. Animals were approximately 3–4 months of age and weighed between 209 and 384 g at the beginning of the experiment. Animals were individually housed in wire-mesh stainless-steel cages or plastic tubs. The vivarium was maintained at an ambient temperature of 21°C and was on a normal 12 h light/dark cycle. All rats were provided ad libitum access to food and water. The sole exception was the rats of the operant self-administration studies, wherein rats were fluid deprived 23 h daily (see below) during the first 2 days of the training phase. Thereafter, these animals were maintained on ad libitum food and water. All training and experimental sessions for all subjects took place between 0900 and 1500.
Operant self-administration studies (systemic and VP): Female P rats (N=35) from the S48 and S49 generations and female HAD-1 rats (N=24) from the S34 generation were used in the present study. No effects of estrous cycle have been observed on drinking patterns in genetically selected rats (McKinzie et al, 1996), and female P rats maintain their body weights within a range that allows for more accurate stereotaxic placement than male P rats (Nowak et al, 1998; June et al, 2001).
Locomotor activity studies: Female P rats (N=42) from the S50 and S51 generations and female HAD-1 rats (N=42) from the S35 generation were used. The treatment of rats for all studies was approved by the institutional review board within the School of Science at IUPUI. In addition, all procedures were conducted in strict adherence with the NIH Guide for the Care and Use of Laboratory Animals.
Drug and Solutions
βCCt
For systemic drug administrations, βCCt was prepared as an emulsion in a Tween-20 solution (Sigma Chemical Co., St Louis, MO) that comprised 100 ml of a 0.90% sodium chloride solution and two drops of Tween-20. βCCt was sonicated (Fisher Scientific, Springfield, NJ) to aid in dissolving the compound. Tween-20 vehicle solution was administered as the control injection for all systemic experiments. Systemic drug injections were given intraperitoneally (i.p.) in an injection volume of 1 ml/kg. For the microinjection studies, βCCt was dissolved in an artificial cerebrospinal fluid (aCSF) (see below).
Other drugs and solutions
EtOH (10% v/v) (USP), saccharin (0.05% w/v), and sucrose (1% w/v) (Fisher Scientific) solutions were prepared for the operant chamber as previously described for oral self-administration (June et al, 1998a, 1998b, 1998c). Chlordiazepoxide was obtained from RBI, (Natick, MA) and mixed in sterile saline (0.9%) for i.p. injections in the locomotor sedation studies. EtOH (10% v/v) was also mixed in sterile saline for i.p. injections in the locomotor sedation studies. A volume sufficient to produce a 1.25 g/kg EtOH dose was employed. The competitive BDZ antagonists ZK 93426 (Schering, Berlin, FRG), flumazenil (Ro15-1788), and the inverse agonist Ro15-4513 (Hoffman La Roche, Nutley, NJ) were donated as gifts for use in the Xenopus oocyte studies.
Operant Self-Administration Procedures
Behavioral testing apparatus
Behavioral testing was conducted in 15 standard operant chambers (Coulbourn Instruments, Allentown, PA) equipped with two removable levers and two dipper fluid delivery systems enclosed in sound-attenuated cubicles as previously described (June et al, 1998a, 1998c). All dipper presentations provided a 1.5-s access to a 0.1-ml solution, followed by a 3-s time out period. Above each lever, three stimulus lights (red, green, and yellow) were present, and a stimulus delivery/reinforcer was indicated by illumination of the middle (green) stimulus light. Responses and reinforcements were recorded and controlled by a Dell computer using the 4.0 Coulbourn L2T2 operant software package (Coulbourn Instruments, Allentown, PA).
Training phase (P rats)
A concurrent fixed-ratio (FR) schedule was employed to investigate the capacity of systemic and direct microinjections of βCCt in the VP to modify EtOH- and saccharin-maintained responses in P rats. The specific details of these procedures have recently been described (June et al, 1998c, 2001). In brief, rats were initially trained to orally self-administered EtOH and water in daily 60-min sessions on a concurrent FR1 schedule in a two-lever choice situation. After a period of stabilization on the FR1 schedule, the response requirement was increased to a concurrent FR4 schedule and the water reinforcer was replaced with saccharin (0.025–0.05% w/v). The importance of alternative and concurrently presented reinforcers in examining the positive reinforcing properties of drugs of abuse has previously been discussed (Meisch and Lemaire, 1993; June, 2002).
Training phase (HAD-1 rats)
To assess the capacity of βCCt to modulate EtOH response in the VP, rats were trained to lever press for alcohol only (see below). However, an alternate-day access paradigm was employed to investigate the capacity of systemic injections of βCCt to modify EtOH- and sucrose-maintained responding. These schedules were employed as previous research has demonstrated that unlike P rats, HAD-1 rats show a profound reduction in EtOH intake (Lankford et al, 1991; Lankford and Myers, 1994) and responding (June, 2002) when presented concurrently with a palatable nondrug reinforcer. In the VP study, rats were initially trained to orally self-administer EtOH and water in daily 60-min sessions on a concurrent FR1 schedule in a two-lever choice situation. After a period of stabilization on the FR1 schedule, the response requirement was increased to a concurrent FR4 schedule and the water reinforcer was gradually eliminated from the protocol and replaced with EtOH. Thus, the VP HAD-1 rats responded concurrently for EtOH solutions on both levers. The VP HAD rats were then stabilized on this regimen for 3 weeks prior to any microinfusions. Responding was considered stable when responses were within ±20% of the average responses for 5 consecutive days.
Rats in the systemic study were trained in a similar manner as the VP animals, except that after a 2-week stabilization period on the concurrently presented EtOH solutions, a series of preliminary studies were conducted to determine the sucrose concentration that produced response rates and profiles similar to that of EtOH during an alternate-day presentation schedule. Following this determination, rats were then stabilized on a regimen of 10% (v/v) EtOH on day 1, and 1% (w/v) sucrose on day 2. This alternate-day paradigm continued for 2 weeks. After this final stabilization period, the drug treatment phase began. Again, responding was considered stable when responses were within ±20% of the average responses for 5 consecutive days. The position of the levers and associated dippers for each reinforcer was alternated daily to control for the establishment of lever preference under all concurrent schedules when two different reinforcers were present.
Systemic drug treatment procedures
βCCt was administered 15 min before the operant session to allow for optimal absorption and CNS distribution. βCCt was tested at doses of 1–40 mg/kg. The duration of the operant sessions was 60 min. Subjects were also tested at 24 and 48 h postdrug administration to determine if any residual drug effects remained with a minimum of 72 and maximum of 96 h allocated between drug treatments to permit animals to return to baseline levels. No problems were encountered in obtaining the βCCt in the Tween-20 solution. This period prevented confounding of drug treatments due to residual effects. The HAD-1 rats were tested at lower doses since our preliminary studies indicated that the dose–response curve was much lower in the HAD line compared with the P rats. All systemic drug treatments were given in a randomized design.
Blood Alcohol Concentration (BAC) Measurement
To ensure that animals were consuming pharmacologically relevant amounts of EtOH during operant sessions, BACs were collected in a subset of animals on days animals did not receive drug treatment. After the first 20 min of an operant session, approximately 100 μl of whole blood was collected from the rats by tail tip into a heparin-coated microsample tube. The BAC samples were collected at the 20-min time point since the majority of EtOH-maintained responsestypically occur during the first half of the operant session (June et al, 2001; Harvey et al, 2002; June, 2002). In addition, an attempt was made to determine the rising phase of the BAC since it has been demonstrated that the rising, but not the falling phase of the blood alcohol curve correlated significantly with the euphoric properties of EtOH (June, 2002). After collection, the whole blood was immediately centrifuged for 5 min at 1100 rpm. Plasma samples of 5 μl were collected with a Gilson Microman M-25 Pipette and injected directly into a GL-5 Analyzer (Analox Instruments; Luxenburg, MA). Microanalysis consisted of measuring the oxygen consumption in the reaction between the sample of alcohol and alcohol oxidase (AOD) using a Clark-type amperometric oxygen electrode. Alcohol reagent buffer solutions (pH 7.4) and alcohol oxidase enzymes were used in all samples tested. Results were calculated in units of mg/dl and printed within 20 s of each trial. To assure accuracy of measurement, each BAC sample was calibrated twice. The mean of the two samples was used as an index of the level of BAC content for a given rat. Single point calibrations were carried out using 5 μl of aqueous 100 mg/dl (21.7 mmol/l) standard.
Surgery
Guide cannulae were stereotaxically implanted bilaterally in the anterior (AP +0.48 mm; ML ±1.6 mm; DV –8.2 mm, with 6° lateral angle) (P rat n=7) and medial (AP –0.26 mm; ML ±2.5 mm; DV –8.0 mm) (P rat n=7; HAD-1 rat n=11) VP according to the Paxinos and Watson (1998) atlas. The neuroanatomical control rats were implanted in either the CPu or NACC. The co-ordinates for the CPu rats (P rat n=3; HAD-1 rat n=4) were AP +1.5; ML ±2.5; DV –4.2. In the NACC group, rats were implanted in the shell (AP +1.4; ML +0.8; DV –6.0) (P rat n=3; HAD-1 rat n=4) or core (AP +1.4; ML ±1.7; DV –5.7) (P rat n=3; HAD-1 rat n=3). Thus, a total of 14 P rats were bilaterally implanted with a cannula in the left and right VP. While a total of nine control P rats were bilaterally implanted with cannulae in the left and right NACC or CPu, a total of 11 HAD-1 rats were unilaterally implanted in the left hemisphere with the guide cannula aimed at the VP, and unilaterally implanted in the right hemisphere with the guide cannula aimed at the CPu/NACC. This strategy was employed to further substantiate the neuroanatomical specificity of the A1 receptor subtype in the ventral striatopallidal area in regulating alcohol-motivated behaviors. In experimental and control animals, the cannulae were aimed 1 mm above the intended brain loci. A stylet that protruded 1 mm beyond the tip of the guide cannulae was inserted when the injector was not in place. The sample sizes obtained after the brains were evaluated under the light microscope for correct cannula placements reflect the data shown in Figures 4 and 7.

Reconstruction of serial coronal sections of the ‘P’ rat brains illustrating the bilateral guide cannula tips for the (a) VP (anterior to posterior division) (N=11) and (b) NACC/Cpu (N=7) (ie neuroanatomical controls). Each rat is represented by two solid black circles: one in the left, and one in the right hemisphere. Coronal sections are adapted from the rat brain atlas of Paxinos and Watson (1998), reproduced with permission from Academic Press.

Reconstruction of serial coronal sections of the HAD-1 rat brains illustrating the unilateral guide cannula tips for the (a) NACC/CPu (n=9) (ie neuroanatomical controls) and (b) VP (anterior to posterior division) (n=9). Each rat is represented by two solid black circles: one in the left NACC/CPu and one in the right VP (total N=9). Coronal sections are adapted from the rat brain atlas of Paxinos and Watson (1998), reproduced with permission from Academic Press.
Microinfusion procedures
The microinfusions were delivered immediately before the operant session with a Harvard infusion pump (Harvard Instruments; South Natick, MA), during which time animals were able to move about freely in their home cages (for details see June et al, 2001). The injection cannula extended 1 mm beyond the tip of the guide cannulae. βCCt was dissolved in aCSF (composition in mM: NaCl, 120; KCl, 4.8; KH2PO4, 1.2; MgSO4, 1.2; NaHCO3, 25; CaCl2, 2.5; D-glucose, 10). When necessary, HCl acid or NaOH was added to the solutions to adjust pH levels to ∼7.4±0.1. βCCt was infused bilaterally in P rats for 5 min at a rate of 0.1 μl/1 min using a 28-gauge injector cannulae. HAD-1 rats were infused unilaterally at a similar rate. Each injector cannula was connected by polyethylene tubing to a 10 -μl Hamilton microsyringe. The injection volume delivered to each hemisphere was 0.5 μl, with a total injection volume for both the left and right hemispheres for both P and HAD-1 rats of 1.0 μl. No problems were encountered in obtaining the βCCt in the aCSF solution. All aCSF and drug treatments were administered in a randomized design in all experiments. P rats received a maximum of seven bilateral infusions, while HAD-1 rats received a maximum of seven unilateral infusions in one hemisphere and seven in the other.
Histology
After the completion of the behavioral testing, animals were killed by CO2 inhalation. Cresyl violet acetate (0.50 μl) was injected into the infusion site, and the brains were removed and frozen. The frozen brains were sliced on a microtome at 50-μm sections and the sections were stained with cresyl violet. Infusion sites were examined under a light microscope and indicated on drawings adapted from the rat brain atlas of Paxinos and Watson (1998). Only rats with correct cannula placements were used in the final data analysis. Three of the P and two of the HAD-1 rats were eliminated due to the right, left, or both cannulas being incorrectly implanted. Additional rats (two from each line) were also eliminated due to inadvertent loss of cannulae in the home cage or operant chamber. A reconstruction of serial coronal sections of the rat brains for P and HAD-1 rats used in the data analysis is depicted in Figures 4 and 7. The coronal sections show that the guide cannulae were implanted in the anterior (Bregma 0.70–0.20 mm) to medial (Bregma −0.26 to −0.30 mm) VP fields, while the control placements were located more dorsally in the NACC and CPu areas.
Statistical analysis
The operant-maintained response data were analyzed by a single-factor repeated measures ANOVA with drug treatment (ie dose) as the independent factor. In the systemic studies, the dependent variables were EtOH- and saccharin-maintained responding in the P rats (N=11) and EtOH- and sucrose-maintained responding for the HAD-1 rats (N=11). In the microinfusion studies, the dependent variables were EtOH- and saccharin-maintained responding for the P rats (VP: N=11; NACC/CPu: N=7) and only EtOH in the HAD-1 rats (VP: N=9; NACC/CPu: N=9). Each dependent variable was analyzed separately. Post hoc comparisons between individual drug treatments were made using the Newman–Keuls test in all experiments. In the concurrent and alternate day schedules, correlated t-tests were used to confirm that response rates for EtOH and saccharin/sucrose responding under baseline and aCSF conditions were similar.
Locomotor Sedation Study
Apparatus
Ambulatory count in the open field was recorded individually for 10 min in a plexiglas chamber (42 cm × 42 cm × 30 cm) using a Digiscan activity monitoring system (Acuscan Electronics, Columbus, Ohio, USA) (for details of the monitoring system, see June et al, 1998b).
Systemic injection procedures
βCCt (15 mg/kg, i.p.) and chlordiazepoxide (10 mg/kg, i.p.) were administered 15 and 30 min, respectively, prior to the rats being placed in the open field. EtOH (1.25 g/kg) was given 5 min prior to placing the rats in the open field. When βCCt was given, in combination with either chlordiazepoxide or EtOH, it was given 10 min prior to chlordiazepoxide and EtOH. As noted above, βCCt was administered in Tween-20 solution, while all other drugs were mixed in sterile saline. Animals were tested between 0900 and 1500.
Statistical analysis
HAD-1 and P rats were randomly assigned to each drug treatment group. Between-group ANOVAs with drug treatment (ie dose) as the independent factor were conducted for both HAD-1 (n=6–9 per treatment group) (total N=42) and P (n=6–8 per treatment group) (total N=42) rats on the locomotor activity parameter (ie ambulatory count). Post hoc comparisons between individual drug treatment groups were made using the Newman–Keuls test in all experiments.
RESULTS
Efficacy of βCCt in Modulating GABA at Recombinant GABAA1–A5 Receptors in Xenopus Oocytes: Comparison with other BDZ Antagonists
βCCt exhibited either a neutral or low-efficacy agonist response at GABAA1 (96±7%) A2 (99±10%), A3 (108±6%), and A4 (107±5%) receptors (Figures 2a). However, a low-efficacy partial inverse agonist response was observed at the A5 receptor (88±7% of the GABA response). Flumazenil exhibited an efficacy profile that was qualitatively similar to βCCt at the A1 (99±5%), A3 (118±7%), and A5 (96±6%) subtypes. At the A2 receptor, flumazenil produced a low-efficacy agonist response (115±4%), while βCCt was GABA neutral (98±10%). Flumazenil also produced a qualitatively similar response to βCCt at the A4 receptor, albeit the magnitude of GABA potentiation by flumazenil far exceeded that of βCCt (132±6 vs 108±6%, respectively). In contrast, ZK 93426 produced a clear agonist profile, potentiating GABAergic activity by 137±8–148±11% across the A1–A4 subtypes, but was GABA neutral at the A5 receptor (96±6%). Figure 2b–j depicts the current traces illustrating the relative magnitude of GABA potentiation by βCCt, flumazenil, and ZK 93426. Despite the qualitatively similar response profile of βCCt and flumazenil, the traces clearly reveal that βCCt did not remarkably affect the GABA currents at the A1, A2, A3, or A4 (data not shown) subtypes relative to the control condition. In contrast, flumazenil significantly increased the GABA currents at the A2 (P<0.09), A3 (P<0.05), and A4 (P<0.01) subtypes relative to the control condition. The traces also confirmed the marked potentiation of the GABA current by ZK 93426 at the A1–A3 and A4 (data not shown) subtypes compared with the control condition (P<0.01).

Modulation of GABAA α1β3γ2, α2β3γ2, α3β3γ2, α4β3γ2, and α5β3γ2 receptor subunit combinations expressed in Ltk cells by βCCt (open bars), flumazenil (shaded bars), and ZK 93426 (black bars). A saturating concentration (1–10 μM) was coapplied over voltage-clamped oocytes along with an EC50 of GABA. (a) Each value is the mean % GABA response±SD of at least four separate oocytes. Actions of βCCt, flumazenil, and ZK 93426 on recombinant GABAA receptor subtypes. Top, current responses of voltage-clamped oocytes expressing GABAA α1β3γ2 receptors (b), during application of 50 μM (EC50) GABA alone for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 50-μM GABA along with 10 μM βCCt for the duration indicated by the open bar (right trace). (c) Current response of a voltage-clamped oocyte during application of 50-μM GABA for the duration indicated by the black bar (left trace). Current response from same oocyte subsequently coapplied with 50-μM GABA along with 1 μM flumazenil for the duration indicated by the open bar (right trace). (d) Current response of a voltage-clamped oocyte during application of 50-μM GABA for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 50-μM GABA along with 10 μM ZK 93426 for the duration indicated by the open bar (right trace). Center, current responses of voltage-clamped oocytes expressing GABAA α2β3γ2 receptors (e), during application of 50 μM (EC50) GABA for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 50-μM GABA along with 10 μM βCCt for the duration indicated by the open bar (right trace). (f) Current response of a voltage-clamped oocyte during application of 50-μM GABA for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 50 μM GABA along with 10 μM flumazenil for the duration indicated by the open bar (right trace). (g) Current response of a voltage-clamped oocyte during application of 50-μM GABA for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 50-μM GABA along with 10 μM ZK 93426 for the duration indicated by the open bar (right trace). Bottom, current responses of voltage-clamped oocytes expressing GABAA α3β3γ2 receptors (h), during application of 30 μM (EC50) GABA for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 30-μM GABA along with 10 μM βCCt for the duration indicated by the open bar (right trace). (i) Current response of a voltage-clamped oocyte during application of 30-μM GABA for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 30-μM GABA along with 1-μM flumazenil for the duration indicated by the open bar (right trace). (j) Current response of a voltage-clamped oocyte during application of 30-μM GABA for the duration indicated by the black bar (left trace). Current response from the same oocyte subsequently coapplied with 30-μM GABA along with 10 μM ZK 93426 for the duration indicated by the open bar (right trace). Scale bars: 5 nA, 10 s.
Neurobehavioral Studies
Blood EtOH content (BAC) determination
Body weights of the P (N=10) and HAD-1 (N=9) rats used for BAC determination ranged from 290 to 407 g. BACs were collected on days that no drug treatments were administered in both rat lines. EtOH responding for P rats yielded intakes of 0.67–2.78 g/kg of absolute EtOH. Consumption in milliliters was 1.45–6.37. BACs ranged from 16 to 92 mg/dl. BACs correlated significantly with EtOH responding (r=0.78, P<0.01) and intake (r=0.82, P<0.01). For the HAD-1 rats, alcohol response yielded intakes of 0.43–1.94 g/kg of absolute EtOH. EtOH consumption in milliliters was 0.84–5.34. BACs ranged from 12 to 86 mg/dl. BACs correlated significantly with EtOH responding (r=0.68, P<0.05) and intake (r=0.66, P<0.05). Once the BACs were determined, group assignments were balanced to include rats from both the low- and high-intake ranges. Thus, rats were semirandomly assigned to their respective groups with a prerequisite being that a given group contained 40–50% of both low or high alcohol responders.
Parenteral Injection Studies
P rats
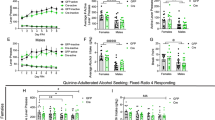
Figure 3a shows that basal operant response rates for EtOH were within 92% of response rates for saccharin (P>0.05). The 5–40 mg/kg βCCt treatments that suppressed responding maintained by alcohol yielded a significant main effect of dose (F(5, 45)=4.64, P<0.01). The Newman–Keuls post hoc tests revealed that all doses significantly suppressed alcohol-maintained responding compared with the control condition (P⩽0.05). At 24 h postdrug administration, the 40 mg/kg dose continued to suppress responding by 76% of control levels (P<0.05). In contrast to the effects on alcohol-maintained responding, βCCt was without effect on responding maintained by saccharin (F(5, 45)=1.64, P>0.05).

Dose–response of systemic βCCt injections (i.p.) in (a) P (Ps) (5–40 mg/kg) and (b) HAD-1 (Hads) (1–10 mg/kg) rats. P rats (N=11) performed under a concurrent FR4 schedule for EtOH (10% v/v) and saccharin (0.05% w/v). HAD-1 rats (N=11) performed under an alternate-day access paradigm, wherein they received EtOH (10% v/v) on day 1 and sucrose (1% w/v) on day 2. At 15 min after the i.p. injections, rats were placed in the operant chamber to lever press for a 60-min session. **P<0.01, *P<0.05 vs the control conditions values by ANOVA and post hoc Newman–Keuls test. Bars represent±SEM in this and subsequent figures.
HAD-1 rats
Figure 3b shows that basal operant responding rates for EtOH and sucrose were very similar (P>0.05). The 1–10 mg/kg βCCt injections dose-dependently suppressed the responding maintained by alcohol, yielding a significant main effect of dose (F(4, 40)=7.84, P<0.001). The Newman–Keuls post hoc tests confirmed that all doses significantly suppressed alcohol-maintained responding compared with the control condition (P⩽0.05). The bottom panel of Figure 3b shows that βCCt suppressed responding maintained by sucrose only with the 10 mg/kg dose; however, the overall ANOVA failed to reach statistical significance (F(4, 40)=1.64, P>0.05). A subsequent post hoc test confirmed that the 10 mg/kg dose produced a marked suppression on responding maintained by sucrose (P<0.01).
Microinfusion Studies
P rats
Figure 4a shows a reconstruction of serial coronal sections of the rat brain illustrating the bilateral guide cannula tips for the 12 VP rats with correct cannula placements. The histological placements show that the guide cannulas were implanted in the dorsal (Bregma 0.70–0.20 mm) to medial (Bregma −0.26 to −0.30 mm) VP fields. Figure 5a–d depict the actual bilateral placements for four of the 12 VP rats in separate photomicrographs illustrating the extent of the lesion sustained as a result of the bilateral guide cannula. The top panel of Figure 6a shows behavioral data for P rats bilaterally infused with βCCt (5–40 μg) compared with the no injection baseline (ie B) and the aCSF control conditions. βCCt dose-dependently reduced EtOH-maintained responding relative to the B and aCSF control conditions resulting in a significant effect of drug dose (F(4, 44)=4.83, P<0.05), (F(4, 44)=5.21, P<0.05, respectively). Post hoc analyses confirmed that all βCCt doses significantly reduced EtOH responding (P⩽0.05). The bottom panel of Figure 6a also shows data for saccharin-maintained responding after the βCCt and control infusions. In contrast to the effects observed on EtOH-maintained responding, βCCt did not alter responding maintained by saccharin with any of the tested doses and produced a nonsignificant effect of drug dose relative to the two control conditions (F(4, 44)=0.98, P>0.05); (F(4, 44)=1.34, P>0.05).

Representative histological photomicrographs of bilaterally implanted cannulas in four ‘P rats’ terminating in the (a) anterior (Bregma +0.70 mm), (b) subcommissural (Bregma +0.20 mm), (c) medial VP (Bregma −0.26 mm), and (d) posterior VP (Bregma −0.80 mm). The photomicrographs depict the distal ends of the cannula tracks.

(a) Performance of female P rats (n=11) on a concurrent FR4 schedule for EtOH (10% v/v) and saccharin (0.05% w/v) following bilateral infusions of βCCt (0.0–40 μg) in the VP. (b) Performance of control female P rats (n=7) on a concurrent FR4 schedule for EtOH (10% v/v) and saccharin (0.05% w/v) following bilateral infusions of βCCt (0.0–40 μg) in the NACC/CPu areas. (c, d) Performance of female Had rats (n=9) on an FR4 schedule for EtOH (10% v/v) following unilateral infusions of βCCt (0.0–7.5 μg) in the VP on the first day of infusion and 24 h postdrug administration. (e) Performance of the same female Had rats in (c) (n=9) on an FR4 schedule for EtOH (10% v/v) following unilateral infusions of βCCt (0.0–7.5 μg) in the NACC/CPu areas on the first day of infusion. **P⩽0.01, *P⩽0.05, compared with the baseline (Bl) and aCSF conditions using post hoc Newman–Keuls tests.
A reconstruction of serial coronal sections for the neuroanatomical control rats is depicted in Figure 4b. The bilateral guide cannula tips for the seven control subjects were at Bregma 2.20 to Bregma 1.20. Figure 6b shows the rates of responding maintained by EtOH (upper panel) and saccharin (lower panel) following bilateral microinjection of the 5–40 μg doses of βCCt. Compared with the aCSF and BL control conditions, none of the βCCt treatments altered EtOH or saccharin-maintained responding. These findings were supported by a nonsignificant effect of drug treatment for EtOH- and saccharin-maintained responding (F(4, 24)= 0.299, P>0.05), (F(4, 24)=1.84, P>0.05), respectively.
HAD-1 rats
As noted above, HAD-1 rats were trained to lever press for alcohol only. In addition, to further substantiate the neuroanatomical specificity of the A1 receptor, HAD-1 rats received a unilateral implant in the VP and a second implant in either the CPu or NACC. Of the 11 rats implanted, nine were implanted in both the VP and CPu/NACC areas. Figure 7a, b show a reconstruction of serial coronal sections for the rats with the correct placements in both loci. The VP cannulae were at Bregma 0.70 to −0.30 (Figure 7a), while the CPu/NACC implants were more dorsal at Bregma 2.20 to −1.20 (Figure 8a). Figures 8a-d and 9a-d, depict representative photomicrographs for four rats, with each having a single cannula track in the CPu/NACC and the other in the anterior to posterior VP. Figure 6c shows the rates of responding maintained by EtOH following unilateral microinjection of the 0.5–7.5 μg doses of βCCt into the VP of HAD-1 rats. Compared with the aCSF control condition, βCCt dose-dependently reduced EtOH response and yielded a significant effect of drug dose (F(5, 40)=4.315, P<0.003). However, post hoc analyses showed that only the 2.5–7.5 μg doses significantly reduced response (P⩽0.05). Figure 6d shows that twenty-four hr postdrug administration, the 2.5–7.5 μg doses continued to reduce responding by as much as 54–63% of control levels (F(5, 40)=4.91, P<0.001). In contrast, Figure 6e shows that unilateral infusions of βCCt into the CPu/NACC areas were completely ineffective in altering alcohol-maintained responding (F(5, 40)=0.466, P>0.05).

Representative histological photomicrographs of two ‘HAD-1 rats’ with one unilateral cannula terminating in the (a, c) NACC/CPu (ie neuroanatomical control loci) and the second unilateral cannula terminating in the (b, d) anterior (Bregma +0.70 mm) to medial VP (Bregma −0.26 mm).

Representative histological photomicrographs of two additional ‘HAD-1 rats’ with one unilateral cannula terminating in the (a, c) NACC/CPu (ie neuroanatomical control loci) and the second unilateral cannula terminating in the (b, d) medial (Bregma −0.26 mm), to posterior VP (Bregma −0.80 mm).
Locomotor Sedation Study: Interaction of βCCt and Chlordiazepoxide
P and HAD-1 rats
Figure 10a, b illustrate the sedative profile of chlordiazepoxide and EtOH. Chlordiazepoxide (10 mg/kg, i.p.) and EtOH (1.25 g/kg, i.p.) produced a profound and comparable reduction in locomotor activity compared with the vehicle-treated controls in the P and HAD-1 rats. These findings resulted in a highly significant effect of drug treatment in P and HAD-1 rats (F(5, 36)= 8.67, P<0.0001) and (F(5, 36)=30.99, P<0.0001), respectively. βCCt (15 mg/kg, i.p.) reversed the sedation produced by both chlordiazepoxide and EtOH in both P (P<0.01 and P<0.01, respectively) and HAD-1 (P<0.01 and P<0.01, respectively) rats. Given alone, βCCt did not produce any intrinsic effects in either rat line (P>0.05).

Evaluation of βCCt's capacity to antagonize the locomotor sedation produced by chlordiazepoxide (CZ) in (a) HAD-1 rats in the vehicle (n=9), 10 mg/kg CZ (n=6), 15 mg/kg βCCt+10 mg/kg CZ (n=7), 1.25 g/kg EtOH (n=7), 15 mg/kg βCCt+1.25g/kg EtOH (n=7), and 15 mg/kg βCCt (n=6) treatment groups and (b) P rats in the vehicle (n=8), 10 mg/kg CZ (n=6), 15 mg/kg βCCt+10 mg/kg CZ (n=7), 1.25g/kg EtOH (n=7), 15 mg/kg βCCt+1.25g/kg EtOH (n=7), and 15 mg/kg βCCt (n=7) treatment groups. Data are ambulatory count in an open field (mean±SEM) for 10 min. **P⩽0.01, compared with the vehicle (Veh) using post hoc Newman–Keuls tests. +P<0.01 compared with the 1.25g/kg EtOH condition. βCCt only partially antagonized the EtOH sedation in P rats.
DISCUSSION
Parenteral administration of βCCt selectively reduced alcohol-maintained responding, but did not alter responding maintained by a highly palatable reinforcer in P rats. In HAD-1 rats, βCCt continued to exhibit selectivity in suppressing alcohol-maintained responding compared with a caloric reward. Further, a four-fold higher dose of βCCt was required to reduce the consumption of the sucrose reinforcer compared to alcohol in this line. Previous research has suggested that when response rates for a drug and alternative reinforcer approximate each other at basal levels, rate is no longer a confounding factor contributing to the effects of an antagonist in drug self-administration studies (Samson et al, 1989; Carroll et al, 1989; Petry and Heyman, 1995; June, 2002). Response rates for EtOH and the alternative reinforcers were similar in P and HAD-1 rats. Hence, βCCt evidenced a marked specificity in reducing alcohol responding compared with responding maintained by other palatable ingesta across two alcohol-preferring lines.
Consistent with the effects observed following parenteral administration, direct VP infusions of βCCt produced dose-dependent reductions in alcohol-maintained responding in both P and HAD-1 rats, but failed to alter responding for a palatable saccharin reward in P rats. These data reinforce the notion that the βCCt-induced reduction of alcohol-maintained behaviors was not due to a general suppression of consummatory behaviors. The βCCt-mediated suppression also exhibited neuroanatomical specificity in P and HAD-1 rats. Hence, suppression was seen at the anterior to posterior VP levels in P and HAD-1 rats, but was not observed with the more dorsal placements in the NACC or CPu in either rat line. Further, this selective typography could clearly be demonstrated even following occupancy of the GABAA1 receptors by βCCt in a single hemisphere. The failure of βCCt to alter alcohol self-administration in the NACC and the CPu could possibly be due to the fact that very few A1 receptors are occupied following a unilateral infusion in the NACC or CPu. However, a more likely explanation for βCCt altering alcohol self-administration is the reported lack of A1 transcript within the NACC and CPu (Churchill et al, 1991; Araki and Tohyama, 1991; Turner et al, 1993; Fritschy and Mohler, 1995; Duncan et al, 1995). These data are also consistent with the marginal levels of [3H]zolpidem binding (a GABAA1 selective agonist) in the NACC and CPu (Duncan et al, 1995). Criswell et al (1993), (1995) and Duncan et al (1995) have suggested that zolpidem binding sites are predictive of loci where EtOH potentiates GABAergic function in the CNS. Finally, recent work in our laboratory has demonstrated that A1 knockout mice consistently fail to initiate lever-press responding for EtOH (10% v/v), while the wild-type mice readily respond for EtOH in the operant chamber (Foster, Homanics, June, unpublished). These data further confirm the significance of the A1 receptor in regulating EtOH-motivated behaviors.
The VP has been reported to play a role in regulating the rewarding properties of both psychostimulant and opioid drugs (Austin and Kalivas, 1990; Hubner and Koob, 1990; Hiroi and White, 1993; Gong et al, 1996, 1997; Johnson and Napier, 1997). However, there has been no direct link of this substrate to the rewarding properties of EtOH. The present study and work with 3-PBC, another GABAA1 agonist-antagonist (Carroll et al, 2000; Harvey et al, 2002), represent the first direct test of the hypothesis. However, it is not known if the GABAA1 receptors of the VP are sufficient to regulate alcohol's reinforcing properties. The VP has a small, albeit much lower density of non-A1 containing GABA receptors (Turner et al, 1993; Fritschy and Möhler, 1995; Wisden et al, 1992). Moreover, the selectivity of βCCt at the GABAA1 receptor compared with the A2 and A3 subtypes is also important to note. Recombinant receptor studies show that βCCt exhibits a >10-fold selectivity for the GABAA1 over the A2 and A3 receptors, and a >110-fold selectivity for the A1 over the A5 subtype (Cox et al, 1995). Thus, binding of βCCt at non-A1 receptors might contribute to the reduction in alcohol responding, even following direct infusion in the VP. However, this hypothesis is mitigated by the failure of βCCt to alter alcohol response in the NACC and CPu, where greater levels of A2 and A3 transcripts have been observed (Turner et al, 1993; Fritschy and Möhler, 1995; Wisden et al, 1992). In addition, GABAergic involvement within the mesolimbic DA or opioid systems in the VP also cannot be ruled out (Austin and Kalivas, 1990; Kalivas et al, 1993). Taken together, the present data do not unequivocally support the role for the GABAA1 receptor as the sole mediator of the antagonistic actions of βCCt, but is the most tenable explanation of the neuromechanisms by which βCCt selectively reduces alcohol responding.
Previous research has demonstrated that parenteral and oral administration of the pyrazoloquinoline (CGS 8216) and the β-carboline (ZK 93426) antagonists can selectively reduce alcohol-maintained response in P rats (June et al, 1998a). At 24 h postdrug administration, the suppression was still detectable with higher doses (⩾20 mg/kg). In contrast, the antagonist flumazenil did not alter EtOH-maintained responding. A single infusion directly into the VP or systemic injection of βCCt was also observed to selectively suppress alcohol responding 24 h postdrug administration under some conditions in P (Figure 3a) and HAD-1 rats (Figure 6d). βCCt's long duration of action makes this compound ideal as a prototype pharmacotherapeutic agent for use with humans. Its longevity in vivo can be attributed to its 3-carboxylate t-butyl ester configuration, which cannot be readily hydrolyzed by esterase activity. t-Butyl ester ligands are apparently too large to fit in the esterase active site and consequently are longer lived in vivo (Zhang et al, 1995).
The locomotor sedation produced by chlordiazepoxide was reversed by βCCt (15 mg/kg, i.p.) in HAD-1 and P rats. These data are consistent with previous research in mutant mice suggesting that the GABAA1 receptor subtype mediates the sedative actions of BDZs (Rudolf et al, 1999; McKernan et al, 2000). However, the current study extends these findings by demonstrating that the GABAA1 receptor may also play a significant role in regulating the sedation produced by an intoxicating dose of alcohol (1.25 g/kg) (Figure 10). These data are also consistent with our previous research demonstrating that ZK 93426 and CGS 8216, but not flumazenil are capable of blocking the sedative actions of alcohol (June et al, 1998c). Hence, given that ZK 93426 and CGS 8216 are nonselective antagonists, it is possible that the βCCt reversal/attenuation of the alcohol-induced sedation may be regulated, in part, by other non-A1 receptor subtypes. Nevertheless, when given alone, βCCt did not produce intrinsic effects on motor activity in either rat line. These data are in agreement with the work of Griebel et al (1999), who reported that doses as high as 60 mg/kg failed to produce intrinsic activity in the open field in mice. Together, the above studies suggest that βCCt is devoid of intrinsic effects on locomotor behaviors and its antagonism of the sedation produced by chlordiazepoxide and alcohol may be mediated via the GABAA1 receptor subtype. Thus, βCCt may be used as a pharmacological tool for distinction among the GABAA receptor subtypes for selected behaviors of alcohol as well as BDZs.
While the behavioral data with βCCt support a role for GABAA1 receptors mediating alcohol's rewarding and sedative properties, the oocyte data demonstrated that even at saturating concentrations, βCCt exhibited a neutral or low-efficacy partial inverse agonist profile at GABAA1, A2 and A5 receptors, and a low-efficacy partial agonist profile at the GABAA3 and A4 subtypes. βCCt was equally, or less potent, than flumazenil in modulating GABAergic activity across the various receptor subtypes. Thus, these data suggest that in the Xenopus oocyte expression system, βCCt displays an efficacy profile characteristic of a neutral BDZ ligand. The functional behavioral significance of such low-level GABAergic modulation is not known. Hence, a clear conundrum in interpreting these data is how can a drug with little or no intrinsic efficacy across the GABAA receptor subtypes modulate alcohol's neurobehavioral effects? One hypothesis is that a low-efficacy BDZ receptor ligand may be sufficient to antagonize alcohol's neurobehavioral effects (see June et al, 2001). This hypothesis has recently received support from our lab by demonstrating that Ro 15-4513 (the most well-studied antialcohol agent) produces a small, albeit statistically significant reduction (∼15%) in GABAergic activity only at recombinant GABAA1 receptor. At the GABAA2–A5 receptors, Ro 15-4513 was GABA neutral (June et al, 2001). Thus, similar to βCCt, Ro 15-4513 exhibits a neutral efficacy across many of the GABAA receptors, but is highly effective in antagonizing several of the neurobehavioral effects of alcohol. A second hypothesis is that efficacy as measured in this Xenopus oocyte assay system does not accurately predict or correlate with the efficacy observations obtained from wild-type receptors, or in whole animal pharmacological experiments. A substantial literature exists on wild-type receptors in the GABA-shift assay (Wong and Skolnick, 1992a, 1992b), TBPS binding (Ticku et al, 1992), and electrophysiological studies (see Harris and Lal, 1988), demonstrating that Ro15-4513 is a partial inverse agonist (also see Suzdak et al, 1986). However, in contrast to the Xenopus oocyte expression system, the above procedures may represent an average efficacy determination of multiple rather than a single subunit isoform (Wong and Skolnick, 1992a, 1992b; Skolnick et al, 1997); hence, these data may not be directly comparable. Finally, subunit combinations other than those used in the present study may be relevant to the effects of βCCt (see Wafford et al, 1993a, 1993b; Barnard et al, 1998; Skolnick et al, 1997).
The fact that βCCt can clearly be differentiated from weak partial inverse agonist such as Ro15-4513 was recently demonstrated in vivo in rodents and primates as well as in vitro in the human HEK cell efficacy assay at the GABAA1, A2 and A3 receptors. For example, in both P and HAD rats, βCCt (5–30 mg/kg) produced anti-anxiety effects in the plus maze comparable to that of chloradiazepoxide (Carroll et al, 2001). Further, βCCt potentiated the anticonflict response produced by an A1 subtype ligands in primates (Paronis et al, 2001). More recently, in the human HEK cell assay we demonstrated that similar to the Xenopus oocyte assay, βCCt displayed weak inverse agonist effects at 0.01–1 μM, however, at higher concentrations (10–100 μM) it produced agonist effects. Interestingly, at the A2, A3, and A4 receptors strong positive GABA modulating effects were observed (41–72%), with the greatest effects being seen at the A2 receptors (Lyddens, June, Cook, unpublished). It should be recalled that the A2 receptor has been proposed to regulate the anxiolytic effects of BDZs (McKernan et al, 2000; Löw et al, 2000). Thus, unlike Ro15-4513, the well-known alcohol antagonist (Suzdak et al, 1986) βCCt, while capable of antagonizing several behavioral effects of alcohol, clearly exhibits GABA agonist effects in several in vivo and in vitro assays.
Based on the present findings, we contend that in contrast to previous reports (for a review see Jackson and Nutt, 1995), a large negative intrinsic efficacy is not a prerequisite for BDZ ligands to antagonize the rewarding or sedative properties of alcohol. Thus, subtype selectivity, as well as efficacy, may be a more important predictor of a BDZ ligand's capacity to selectively antagonize alcohol's neurobehavioral actions in the absence of intrinsic effects (June et al, 2001). In further support of this hypothesis, molecular modeling studies have shown that the pharmacophore for high affinity and selectivity at the GABAA1 receptor is clearly different for βCCt compared to the prototypical BDZ antagonist flumazenil (Cox et al, 1998).
In conclusion, the present study demonstrated that in two rodent models of chronic alcohol consumption (P and HAD-1 lines), βCCt produces reliable and selective antagonism of EtOH-seeking behaviors. We further demonstrated that the βCCt suppression was regulated in part, via the VP, a mesolimbic substrate purported to play a role in the reinforcing properties of other abuse drugs (Hubner and Koob, 1990; Gong et al, 1996; Johnson and Napier, 1997). βCCt's selectivity at the GABAA1 receptor and low intrinsic efficacy (close to GABA neutral) across the various receptor subtypes may contribute to its failure to exhibit locomotor-impairing effects in the current study. It has been suggested that greater side effects of BDZs occur with ligands that lack receptor subtype selectivity (Stephens et al, 1992). However, the degree to which the oocyte data accurately reflects βCCt's in vivo actions remains to be determined. Nevertheless, we conclude that βCCt may have potential as a prototype pharmacotherapeutic agent to effectively reduce alcohol drinking behavior in human alcoholics. βCCt's capacity to reduce alcohol's euphorigenic properties, while concurrently eliminating or attenuating its motor-impairing effects, should render it an optimal prototype in the development of pharmacotherapeutic agents to treat alcohol-dependent individuals.
References
Araki T, Tohyama M (1991). Region-specific expression of GABAA receptor α3 and α4 subunits mRNAs in the rat brain. Mol Brain Res 12: 295–314.
Austin MC, Kalivas PW (1990). Enkephalinergic and GABAergic modulation of motor activity in the ventral pallidum. J Pharmacol Exp Ther 252: 1370–1377.
Barnard EA, Skolnick P, Olsen RW, Möhler W, Sieghart W, Biggio G et al (1998). International union of pharmacology. XV. Subtypes of γ-aminobutyric acid receptors: classification on the basic of subunit structure and receptor function. Pharmacol Rev 50: 291–313.
Carroll M, Foster KL, Harvey S, Mc Kay PF, Cook JM, June HL (2000). Selective GABAA-α1 subunit ligands (βCCt, 3-PBC) attenuate responding maintained by ethanol following microinjection into the ventral pallidum. Alcoholism: Clin Exp Res 24: 47A.
Carroll ME, Lac ST, Nygaard SL (1989). A concurrently available nondrug reinforcer prevents the acquisition or decreases the maintenance of cocaine-reinforced behaviors. Psychopharmacology 97: 23–29.
Carroll M, Woods II JE, Seyoum RA, June HL (2001). The role of the GABAa α1 subunit in mediating the sedative and anxiolytic properties of benzodiazepines. Alcoholism: Clin Exp Res 25: 12A.
Churchill L, Bourdelais A, Austin S, Lolait SJ, Mahan LC, O'Carroll AM et al (1991). GABAA receptors containing α1 and β2 subunits are mainly localized on neurons in the ventral pallidum. Synapse 8: 75–85.
Churchill L, Kalivas PW (1994). A topographical organized GABA projection from the ventral pallidum to the nucleus accumbens in the rat. J Comp Neurol 345: 579–595.
Cloninger R (1987). Neurogenetic adaptive mechanisms in Alcoholism. Science 236: 410–416.
Cox ED, Diaz-Arauzo H, Huang Q, Mundla SR, Chunrong M, Harris B et al (1998). Synthesis and evaluation of analogues of the partial agonist 6-(Propyloxy)-4-(methoxymethyl)-β-carboline-3-carboxylic acid ethyl ester (6-PBC) and the full agonist 6-(Benzyloxy)-4-(methoxymethyl)-β-carboline-3-carboxylic acid ethyl ester (ZK 93423) at wild type and recombinant GABAA receptors. J Med Chem 41: 2537–2552.
Cox ED, Hagen TJ, McKernan RM, Cook JM (1995). BZ1 receptor specific ligands. Synthesis and biological properties of BCCt, A BZ1 receptor subtype specific antagonist. Med Chem Res 5: 710–718.
Criswell HE, Simson PE, Duncan GE, Mc Cown TJ, Herbert JS, Morrow AL et al (1993). Molecular basis for regionally specific action of ethanol on γ-aminobutyric acidA receptors: generalization to other ligand-gated ion channels. J Pharmacol Exp Ther 267: 522–527.
Criswell HE, Simson PE, Knapp DJ, Devaud LL, Mc Cown TJ, Duncan GE et al (1995). Effect of zolpidem on γ-aminobutyric acid (GABA)-induced inhibition predicts the interaction of ethanol with GABA on individual neurons in several rat brains. J Pharmacol Exp Ther 273: 526–536.
DSM-IV-TR (2000). Diagnostic and Statistical Manual of Mental Disorders, 4th Edition, Text Revision. American Psychiatric Association. Alcohol Related Disorders, pp 212–223.
Duncan GE, Breese GR, Criswell HE, McCown TJ, Herbert JS, Devaud LL et al (1995). Distribution of {3Hzolpidem binding sites in relation to messenger RNA encoding the α1, β2 and γ2 Subunits of GABAA receptors in rat brain. Neuroscience 64: 1113–1128.
Fritschy JM, Mohler H (1995). GABAA-receptor heterogeneity in the adult rat brain. Differential regional and cellular distribution of seven major subunits. J Comp Neurol 359: 154–194.
Gong W, Justice JB, Neill D (1997). Dissociation of locomotor and conditioned place preference responses following manipulation of GABA-A and AMPA receptors in ventral pallidum. Prog Neuropsychopharmacol Biol Psychiatry 21: 839–852.
Gong W, Neill D, Justice JB (1996). Conditioned place preference and locomotor activation produced by injection of psychomotor stimulants in the ventral pallidum. Brain Res 707: 64–74.
Griebel G, Perrault G, Letang V, Grainger P, Avenet P, Schoemaker H et al (1999). New evidence that the pharmacological effects of benzodiazepine receptor ligands can be associated with activities at different BZ (α) receptor subtypes. Psychopharmacology (Berl) 146: 205–213.
Groenewegen HJ, Berende HW, Haber SN (1993). Organization of the output of the ventral striatopallidal system in the rat: ventral pallidal efferents. Neuroscience 57: 113–142.
Harris CM, Lal H (1988). Central nervous system effects of Ro15-4513. Drug Dev Res 13: 187–203.
Harvey SC, Foster KL, McKay PF, Carroll M, Seyoum R, Woods II JE et al (2002). The GABAA receptor α1 subtype in the ventral pallidum regulates EtOH-seeking behaviors. J Neurosci 22: 3765–3775.
Harvey SC, McIntosh JM, Cartier GE, Maddox FN (1997). Determinants of alpha-conotoxin MII on α3β2 neuronal nicotinic receptors. Mol Pharmacol 51: 336–342.
Heimer L, Alheid G (1991). Piecing together the puzzle of basal forebrain anatomy. In: Napier TC, Kalivas PW, Hanin I (eds) The Basal Forebrain: Anatomy to Function. Plenum Press: New York. pp 1–42.
Heimer L, Zahm DS, Churchill P, Kalivas W, Wohltmann C (1991). Specificity in the projection patterns of accumbal core and shell in the rat. Neuroscience 41: 89–125.
Hiroi N, White NM (1993). The ventral pallidum area is involved in the acquisition but not the expression of the amphetamine conditioned place preference. Neurosci Lett 156: 912.
Hubner CB, Koob GF (1990). The ventral pallidum plays a role in mediating cocaine and heroin self-administration in the rat. Brain Res 508: 20–29.
Jackson HC, Nutt DJ (1995). Inverse agonist and alcohol. In: Sarter M, Nutt DJ, Lister RG (eds) Benzodiazepine Receptor Inverse Agonists. Wiley-Liss: New York. pp 243–270.
Johnson BA, Ait-Daoud N (2000). Neuropharmacological treatments for alcoholism: scientific basis and clinical findings. Psychopharmacology (Berl) 149: 327–344.
Johnson PI, Napier TC (1997). Morphine modulation of GABA and glutamate-induced changes of ventral pallidal neuronal activity. Neuroscience 77: 187–197.
June HL (2002). Preclinical models to evaluate potential pharmacotherapeutic agents in treating alcoholism and studying the neuropharmacological bases of alcohol-seeking behaviors. In: J Crawley, C Gerfen, R McKay, M Rogawski, D Sibley, P Skolnick (eds) Current Protocols in Neuroscience Vol. 9 John Wiley & Sons, Inc.: New York, NY. pp 1–23.
June HL, Dejaravu SL, Williams J, Cason CR, Eggers MW, Greene TL et al (1998b). GABAergic modulation of the behavioral actions of ethanol in alcohol-preferring (P) and nonpreferring (NP) rats. Eur J Pharmacol 342: 139–151.
June HL, Grey C, Warren-Reese C, Lawrence A, Thomas A, Cummings R et al (1998c). The opioid receptor antagonist nalmefene reduces alcohol motivated behaviors: preclinical studies in alcohol preferring (P) and outbred Wistar rats. Alcoholism: Clin Exp Res 22: 2174–2185.
June HL, Harvey SC, Foster KL, McKay PF, Cummings R, Garcia M et al (2001). GABAA-receptors containing α5 subunits in the CA1 and CA3 hippocampal fields regulate ethanol-motivated behaviors: an extended ethanol reward circuitry. J Neurosci 21: 2166–2177.
June HL, Zucarelli D, Craig KS, DeLong J, Cason CR, Torres L et al (1998a). High affinity benzodiazepine antagonists reduce responding maintained by EtOH presentation in ethanol-preferring (P) rats. J Pharmacol Exp Ther 284: 1006–1014.
Kalivas PW, Churchill L, Klitenick MA (1993). GABA and enkephalin projection from the nucleus accumbens and ventral pallidum to the ventral tegmental area. Neuroscience 57: 1047–1060.
Kessler RC, Crum RM, Warner LA (1997). The lifetime co-occurrence of DSM-III-R alcohol abuse and dependence with other psychiatric disorders in the National Comorbidity Survey. Arch Gen Psychiatry 54: 313–321.
Koob GF (1998). The role of the striatopallidal and extended amygdala systems in drug addiction. Ann NY Acad Sci 877: 445–460.
Koob GF, Roberts AJ, Shulties G, Parsons LH, Heyser CJ, Hyytia P et al (1998). Neurocircuitry targets in ethanol reward and dependence. Alcoholism: Clin Exp Res 22: 3–9.
Kranzler HR (2000). Pharmacotherapy of alcoholism: gaps in knowledge ad opportunities for research. Alcohol Alcoholism 35: 537–547.
Kuo H, Chang HT (1992). Ventral-pallidostriatal pathway in the rat brain. A light electron microscopic study. J Comp Neurol 321: 626–636.
Lankford MF, Myers RD (1994). Genetics of alcoholism: simultaneous presentation of a chocolate drink diminishes alcohol preference in high alcohol drinking rats. Pharmacol Biochem Behav 49: 417–425.
Lankford MF, Roscoe AK, Pennington SN, Meyers RD (1991). Drinking of high concentrations of ethanol vs palatable fluids in alcohol-preferring (P) rats: valid animal model of alcoholism. Alcohol 8: 293–299.
Li T-K (2000). Pharmacogenetics of responses to alcohol and genes that influence alcohol drinking. J Stud Alcohol 61: 5–12.
Li T-K, Crabb DW, Lumeng L (1991). Molecular and genetic approaches to understanding alcohol-seeking behavior. In: Meyer RE, Koob GF, Lewis MJ, Paul SP (eds) Neuropharmacol Ethanol. Birkhauser: Boston. pp 107–124.
Löw K, Crestani F, Keist R, Benke D, Brünig I, Benson JA et al (2000). Molecular and neuronal substrate for the selective attenuation of anxiety. Science 290: 131–134.
Lumeng L, Murphy JM, McBride WJ, Li T-K (1995). Genetic influences on alcohol preference in animals. In: Begleiter H, Kissin B (eds) The Genetics of Alcoholism. Oxford University Press: New York. pp 165–201.
McBride WJ, Li T (1998). Animal models of alcoholism: neurobiology of high alcohol-drinking behavior in rodents. Crit Rev Neurobiol 12: 339–369.
McKernan RM, Rosh TW, Reynolds DS, Sure C, Wafford KA, Attack JR et al (2000). Sedative but not anxiolytic properties of benzodiazepines are mediated by the GABAA receptor α1 subtype. Nat Neurosci 3: 587–592.
McKinzie DL, Eha R, Murphy JM, McBride WJ, Lumeng L, Li T-K (1996). Effects of taste aversion training on the acquisition of alcohol drinking in adolescent P and HAD rat lines. Alcoholism: Clin Exp Res 20: 682–687.
Meisch RA, Lemaire GA (1993). Drug self-administration. In: Van Haren F (ed) Methods in Behavioral Pharmacology. Elsevier: New York. pp 257–300.
Mogenson GJ, Nielson MA (1983). Evidence that an accumbens to subpallidal GABAergic projection contributes to locomotor activity. Brain Res Bull 11: 309–314.
Napier TC, Chrobak JJ (1992). Evaluation of ventral pallidum dopamine receptor activation in behaving rats. Neuroreport 3: 609–611.
National Institute of Alcohol Abuse and Alcoholism (NIAAA) (1997). Ninth Special Report to the US Congress on Alcohol and Health, NIH Pub. No 97-4017. National Institute on Alcohol and Alcoholism: Bethesda, MD.
Nauta WJH, Smith GP, Faull RLM, Domesick VB (1978). Efferent connections and nigral afferents of the nucleus accumbens septi in the rat. Neuroscience 3: 385–401.
Nowak KL, McBride WJ, Lumeng L, Li T-K, Murphy JM (1998). Blocking GABAA receptors in the anterior ventral tegmental area attenuates ethanol intake of the alcohol-preferring P rat. Psychopharmacology (Berl) 139: 108–116.
O'Malley SS, Jaffe AJ, Chang G, Schottenfeld RS, Meyer RE, Rounsaville B (1992). Naltrexone and coping skills therapy for alcohol dependence: a controlled study. Arch Gen Psychiatry 49: 881–887.
Paronis CA, Cox ED, Cook JM, Bergman J (2001). Different types of GABAa receptors may mediate the anticonflict and response rate-decreasing effects of zalepon, zolpidem, and midazolam in squirrel monkeys. Psychopharmacology (Berl) 156: 461–468.
Paxinos G, Watson C (1998). The Rat Brain in Stereotaxic Coordinates. Academic Press: Sydney.
Petry NM, Heyman GM (1995). Behavioral economic analysis of concurrent ethanol/sucrose and sucrose reinforcement in the rat: effects of altering variable ratio requirements. J Exp Anal Behav 64: 331–359.
Rowlett JK, Tornatzky W, Cook JM, Chunrong M, Miczek KA (2001). Zolpidem, triazolam, and diazepam decrease distress vocalizations in mouse pups: differential antagonism by flumazenil and the β-carboline-3-carbozylate-t-butyl ester (βCCt). J Pharmacol Exp Ther 297: 247–253.
Rudolf U, Crestani F, Benke D, Brünig I, Benson JA, Fritschy JM et al (1999). Benzodiazepine actions mediated by specific γ-aminobutyric acidA receptor subtypes. Nature 401: 796–800.
Samson HH, Haraguchi M, Tolliver GA, Sadeghi KG (1989). Antagonism of ethanol-reinforced behavior by the benzodiazepine inverse agonists R015-4513 and FG 7142: relationship to sucrose reinforcement. Pharmacol Biochem Behav 33: 601–608.
Sanger DJ, Benavides J, Perrault G, Morel E, Cohen C, Joly D et al (1994). Recent developments in the behavioral pharmacology of benzodiazepine (omega) receptors: evidence for the functional significance of receptor subtypes. Neurosci Biobehavi Rev 18: 355–372.
Shannon HE, Gruzman F, Cook JM (1984). β-carboline-3-carboxylate-t-butyl ester: a selective BZ1 benzodiazepine receptor antagonist. Life Sci 35: 2227–2236.
Skolnick P, Hu RJ, Cook CM, Hurt SD, Trometer JD, Lui R et al (1997). [3H] RY 80: a high affinity, selective ligand for γ-aminobutyric acidA receptors containing alpha-5 subunits. J Pharmacol Exp Ther 283: 488–493.
Spanagel R, Zieglgansberger T (1997). Anti-craving compounds for ethanol: new pharmacological tool to study addictive processes. Trends Pharmacol Sci 18: 54–59.
Stephens DN, Turski L, Hillman M, Turner JD, Schneider HH, Yamaguchi M (1992). What are the differences between abecarnil and conventional benzodiazepine anxiolytics. In: Biggio G, Concas A, Costa E (eds) GABAergic Synaptic Transmission (Molecular, Pharmacological, and Clinical Aspects). Advances in Biochemical Pharmacology. Raven Press: New York. pp 395–405.
Suzdak P, Glowa JR, Crawley JN, Schwartz RD, Skolnick P, Paul SM (1986). A selective imidazodiazepineantagonist of ethanol in the rat. Science 234: 1243–1247.
Ticku MK, Mhatre M, Mehta AK (1992). Modulation of GABAergic transmission by alcohol. In: Biggio G, Concas A, Costa E (eds) GABAergic Synaptic Transmission (Molecular, Pharmacological, and Clinical Aspects). Advances in Biochemical Pharmacology. Raven Press: New York. pp 255–267.
Turner JD, Bodewitz G, Thompson CL, Stephenson FA (1993). Immunohistochemical mapping of gamma-aminobutyric acid type-A receptor alpha subunits in rat central nervous system. In: DN Stephens (ed) Anxiolytic β-carbolines: From Molecular Biology to the Clinic. Springer-Verlag: New York. pp 29–49.
Volpicelli JR, Alterman AI, Hayashida M, O'brien CP (1992). Naltrexone and the treatment of alcohol dependence. Arch Gen Psychiatry 49: 876–880.
Wafford KA, Bain CJ, Whiting PJ, Kemp JA (1993a). Functional comparison of the role of γ subunits in recombinant human γ-aminobutyric acidA/benzodiazepine receptors. Mol Pharmacol 44: 437–442.
Wafford KA, Whiting PJ, Kemp JA (1993b). Differences in affinity of benzodiazepine receptor ligands at recombinant γ-aminobutyric acidA receptors subtypes. Mol Pharmacol 43: 240–244.
Wisden H, Laurie DJ, Monyer H, Seeburg PH (1992). The distribution of 13 GABAA receptors subunit mRNAs in the rat brain: telencephalon, diencephalon, mesencephalon. J Neurosci 12: 1040–1062.
Wong G, Skolnick P (1992a). High affinity ligands for ‘diazepam insensitive’ benzodiazepine receptors. Eur J Pharmacol: Mol Pharmacol Sect 225: 63–68.
Wong G, Skolnick P (1992b). Ro15-4513 binding to GABAA receptors: subunit composition determines ligand efficacy. Pharmacol Biochem Behav 42: 107–110.
Zahm DS, Heimer L (1988). Ventral striatopallidal parts of the basal ganglia in the rat. I. Neurochemical compartmentation as reflected by the distributions of neurotensin and substance P immunoreactivity. J Comp Neurol 272: 516–535.
Zhang P, Koehler KF, Cook JM (1995). Development of a comprehensive pharmacophore model for the benzodiazepine receptor. Drug Design Discovery 12: 193–248.
Acknowledgements
We thank Drs T-K Li and Larry Lumeng and the Alcohol Research Center at Indiana University School of Medicine for providing the P and HAD-1 rats. This research was supported in part by Grants AA10406 and AA11555 (HLJ) from the National Institute of Alcohol Abuse and Alcoholism and MH 46851 (JMC) from the National Institute of Mental Health (NIH). Katrina L Foster, Cecily M Jones, and Marin Garcia were supported in part by Grant T35M from the National Heart, Lung, and Blood Institute of NIH (Short Term Training Program for Minority Students in Biomedical Research). Katrina Foster was also supported in part by a Minority Neuroscience Fellowship from the American Psychological Association. We also thank Schering (Berlin, FRG) for their generous donation of ZK 93426 and Hoffman La Roche (Nutley, NJ) for their donation of flumazenil (Ro15-1788). Finally, we thank Drs Luddens (University of Mainz, Mainz, Germany) and Mahan (National Institutes of Health) for supplying some of the cDNA clones.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
June, H., Foster, K., McKay, P. et al. The Reinforcing Properties of Alcohol are Mediated by GABAA1 Receptors in the Ventral Pallidum. Neuropsychopharmacol 28, 2124–2137 (2003). https://doi.org/10.1038/sj.npp.1300239
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1300239
Keywords
This article is cited by
-
Lateral preoptic and ventral pallidal roles in locomotion and other movements
Brain Structure and Function (2018)
-
Comparison of the locomotor-activating effects of bicuculline infusions into the preoptic area and ventral pallidum
Brain Structure and Function (2014)
-
Effects of the benzodiazepine GABAA α1-preferring ligand, 3-propoxy-β-carboline hydrochloride (3-PBC), on alcohol seeking and self-administration in baboons
Psychopharmacology (2013)
-
Role for ventral pallidal GABAergic mechanisms in the regulation of ethanol self-administration
Psychopharmacology (2012)
-
Neurocircuitry of Addiction
Neuropsychopharmacology (2010)
