
Abstract
Teenage drinking is a cause of growing concern in industrialized countries, where almost 35% of alcohol drinkers are under 16 years old. Increased anxiety, irritability, and depression among adolescents may induce them to seek the anxiolytic and rewarding properties of alcohol. We studied the effects of acute alcohol intoxication (AAI) on the plasma levels of progesterone and allopregnanolone in female adolescents. Blood samples were drawn from female adolescents who arrived at the emergency department. One study group was formed by those who arrived with evident behavioral symptoms of AAI and the other by those arriving for mild trauma (contusions, sprains) after no consumption of alcohol (controls). Our results demonstrate that AAI significantly increases serum progesterone and allopregnanolone levels in both follicular and luteal phases of the ovarian cycle. Since alcohol and allopregnanolone positively modulate gamma-aminobutyric acid type A (GABAA) receptors, allopregnanolone may play a major role in the anxiolytic and rewarding effects of alcohol, either directly or by influencing the sensitivity of GABAA-receptors to alcohol.
Similar content being viewed by others

INTRODUCTION
Alcohol abuse among adolescents is a widespread and growing threat to the health of young people in industrialized countries (Bailey and Valery, 1993), where almost 35% of habitual drinkers are under 16 years old (Shope et al, 1994). A growing number of young adolescents drink alcoholic beverages every weekend.
Our group has reported the deleterious effects of alcohol abuse on the reproductive and growth axis of adolescents (Frias et al, 2000a,2000b). An important question to be addressed is why increasing numbers of adolescents consume alcohol, despite the adverse and probably long-lasting effects of this drug on their behavior and physiology.
Owing to the pace of life in modern industrialized society, among other factors, adolescents may feel that their affective needs are not fully met, leading to the development of symptoms of anxiety, irritability, and depression. This could in part explain why these young people seek the anxiolytic and rewarding properties of alcohol, a socially permitted drug.
Alcohol positively modulates gamma-aminobutyric acid type A (GABAA) receptors and produces sedative and anxiolytic effects. There is evidence for interaction between alcohol and neurosteroids at GABAA-receptors (Grobin et al, 1998). The neurosteroid allopregnanolone is one of the most potent endogenous positive modulators of GABAA-receptors and has sedative, hypnotic, anticonvulsant, and anxiolytic effects (Lambert et al, 1995). In animals, it has been suggested that allopregnanolone produced in response to systemic ethanol administration could contribute to several of the effects of ethanol that are associated with the modulation of GABAA-receptors (Morrow et al, 2001a). Neurosteroids have been shown to be rewarding in rodents (Finn et al, 1997). Therefore, neurosteroids may contribute to the anxiolytic and rewarding effects of alcohol.
Many animal studies have investigated the effect of alcohol on neurosteroid concentrations (Barbaccia et al, 1999). However, we know of no studies that have demonstrated the effect of alcohol on neurosteroids in adolescent humans, possibly because of the difficulty to recruit appropriate study subjects. Studies involving the controlled administration of alcohol are evidently ruled out in adolescents on ethical grounds.
The present study aimed to determine allopregnanolone levels in female adolescent humans in situations of acute alcohol intoxication (AAI).
MATERIALS AND METHODS
A study was conducted of 35 white female adolescents (13–17 years old) of variable weight, height, and educational level who arrived at the emergency department of our hospital between 12.00 and 3.30 am. The subjects were assigned to one of the two study groups. One group (AAI, n=20) was formed by those arriving at the emergency department with evident behavioral symptoms of AAI (slurred speech, unstable walking); after remission of these symptoms, all of them reported that they habitually became drunk every weekend. The second group (controls, n=15) comprised young women who arrived at the department for mild trauma (contusions, sprains) and reported no alcohol consumption. All subjects were interviewed regarding their current ovarian phase and any drug treatments, in the case of AAI after the remission of their symptoms. No subjects were receiving medication at the time of the study or suffered any apparent endocrine disorder. All participants, or their parents, gave their informed consent for participation in the study, which was carried out in accordance with the Helsinki Declaration of 1975.
Blood samples were extracted from all subjects on arrival at the emergency area. Serum samples were frozen and stored at −20°C. Assessment of progesterone concentrations was performed by radioimmune assay (RIA), using commercially available Sorin Biomedica kits (Vercelli, Italy) without modifications. Allopregnanolone was measured by RIA as previously described (Torres et al, 2001), using an antiserum kindly provided by Roussel–Uclaf. The plasma samples were extracted with diethyl ether (Genazzani et al, 1998) and the extracts were purified according to Corpechot et al (1993). Since the ovary is an important source of circulating progesterone and allopregnanolone in humans (Rapkin et al, 1997; Genazzani et al, 2002), the study included women in follicular (AAI, n=10; controls, n=7) or luteal (AAI, n=10; controls, n=8) phases of their ovarian cycle. Their consumption of alcohol was confirmed by the measurement of blood alcohol concentrations, using a gas-chromatographic head-space method described previously by our group (Frias et al, 2002).
Statistically significant differences between the groups were analyzed by a two-way ANOVA. Since the Levene test for equality of variance showed significance, logarithmic transformation was performed, meeting the assumption of homogeneity of variance. The Bonferroni method was used in this study. The SPSS version 9.0 for Windows software package was used in the statistical analysis. Results are expressed as mean±SEM.
RESULTS
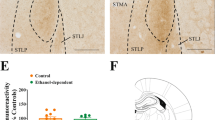
Serum levels of allopregnanolone and progesterone in the AAI and control groups, respectively, are shown in Figures 1 and 2. Two-way ANOVA of the allopregnanolone results showed significantly higher allopregnanolone levels in the AAI group vs controls (F1,31=75.33, P<0.001) and significantly higher allopregnanolone levels in luteal vs follicular phase (F1,31=47.36, P<0.001). Application of the Bonferroni method showed that the AAI females had significantly higher allopregnanolone levels vs controls in both follicular and luteal phase (P<0.01). With respect to progesterone values, significantly higher levels were found in AAI women vs controls (F1,31=59.14, P<0.001), and progesterone levels were significantly higher in luteal than follicular phase (F1,31=770.07, P<0.001). Application of the Bonferroni method showed that progesterone levels were significantly higher in AAI women vs controls in both follicular and luteal phase (P<0.01).

Serum allopregnanolone in AAI and controls (C). The data are expressed as mean±SEM. The Bonferroni method showed statistical differences in AAI vs C P<0.01 in both follicular and luteal phases of the ovarian cycle.

Serum progesterone in AAI and controls (C). The data are expressed as mean±SEM. The Bonferroni method showed statistical differences in AAI vs C P<0.01 in both follicular and luteal phases of the ovarian cycle.
There was no significant difference in the blood alcohol level of the AAI adolescents between follicular and luteal phases (108±4 vs 99±3.5 mg/dl, respectively).
DISCUSSION
According to the present results, AAI raises the plasma concentrations of progesterone and allopregnanolone in adolescent females during both the luteal and follicular phases of their ovarian cycle. These findings are consistent with a previous report that alcohol injection increased plasma concentrations of allopregnanolone in rats (Barbaccia et al, 1999). Although the effects of ethanol on the hepatic metabolism of steroids should be taken into account, our data may indicate that alcohol ingestion promotes neurosteroidogenesis in peripheral organs. A previous study reported that exogenously administered allopregnanolone attenuated the stress-induced release of corticotropin-releasing factor (CRF), adrenocorticotropin hormone (ACTH), and corticosterone and blunted the anxiogenic effect of CRF. Our group earlier reported that AAI produces a high increase in pituitary–adrenal axis hormones in female adolescents (Frias et al, 2000a). Circulating allopregnanolone is increased in humans and rats after CRF and ACTH administration (Genazzani et al, 1998; Torres et al, 2001). Allopregnanolone may therefore represent a novel feedback of the hypothalamic–pituitary–adrenal axis, as previously proposed (Purdy et al, 1991).
The ovary and adrenal cortex are sources of circulating allopregnanolone. Although we could not know the contribution of each source, we observed an increase in serum allopregnanolone in our AAI females in both follicular and luteal phase. Earlier reports demonstrated that circulating allopregnanolone crosses the blood–brain barrier (Morrow et al, 2001b) and may exert its anxiolytic, analgesic, anticonflictive, and rewarding effects in the CNS through allosteric modulation of GABAA-receptors.
The increase in allopregnanolone in response to AAI in female adolescents may have major biological repercussions. Allopregnanolone may increase the anxiolytic or rewarding effects of this socially permitted drug either directly, or by increasing the sensitivity of GABA receptors to alcohol.
References
Bailey SL, Valery RJ (1993). Dimensions of adolescent problem drinking. J Stud Alcohol 54: 555–565.
Barbaccia ML, Affricano D, Trabucchi M, Purdy RH, Colombo G, Agabio R et al (1999). Ethanol markedly increases ‘GABAergic’ neurosteroids in alcohol-preferring rats. Eur J Pharmacol 384: R1–R2.
Corpechot C, Young J, Calvel M, Wehrey C, Veeeltz N, Touyer G et al (1993). Neurosteroids 3α Hydroxi-5α Pregnan-20-one and its precursors in the brain, plasma and steroidogenic gland of male and female rats. Endocrinology 133: 1003–1009.
Finn DA, Phillip TJ, Okorn DM, Chester JA, Cunningham CL (1997). Rewarding effects of the neuractive steroid 3α-hydroxy-5α-pregnan-20-one in mice. Pharmacol Biochem Behav 56: 261–264.
Frias J, Rodriguez R, Torres JM, Ruiz E, Ortega E (2000a). Effects of acute alcohol intoxication on pituitary–gonadal axis, pituitary–adrenal axis hormones β-endorphin and prolactin in human adolescents of both sexes. Life Sci 67: 1081–1806.
Frias J, Torres JM, Miranda MT, Ruiz E, Ortega E (2002). Effects of acute alcohol intoxication on pituitary–gonadal axis, pituitary–adrenal axis hormones β-endorphin and prolactin in human adults of both sexes. Alcohol Alcoholism 37/2: 169–173.
Frias J, Torres JM, Rodriguez R, Ruiz E, Ortega E (2000b). Effects of acute alcohol intoxication on growth axis in human adolescents of both sexes. Life Sci 67: 2691–2697.
Genazzani AD, Luisi M, Malavasi B, Stucchi C, Stefano L, Casarosa E et al (2002). Pulssatile secretory characteristics of allopregnanolone, a neuroactive steroids, during the menstrual cycle and in amenorrheic subjects. Eur J Endocrinol 146: 347–356.
Genazzani AR, Petraglia F, Bernardi F, Casarosa E, Salvestroni C, Tonetti A et al (1998). Circularing levels of allopregnanolone in humans: gender, age and endocrine influences. J Clin Endocr Metab 83: 2099–2103.
Grobin AC, Matthees DB, Devaud LL, Morrow AL (1998). The role of GABAA receptors in the acute and chronic effects of ethanol. Psychopharmacology 139: 2–19.
Lambert JJ, Belelli D, Hill-Venning C, Peters JA (1995). Neurosteroids and GABA receptors function. Trend Pharmacol Sci 16: 295–303.
Morrow AL, VanDoren MJ, Fleming R, Penland S (2001b). Ethanol and neurosteroid interactions in the brain. Int Rev Neurobiol 46: 349–377.
Morrow AL, VanDoren MJ, Penland SN, Matthew DB (2001a). The role of GABAergic neuroactive steroids in ethanol action, tolerance and dependence. Brain Res Brain Res Rev 37: 98–109.
Purdy RH, Morrow L, Moor PH, Paul SM (1991). Stress-induced elevation of aminobutiricacid type A receptor active steroids in the brain. Proc Natl Acad Sci USA 88: 4553–4557.
Rapkin AJ, Morgan M, Goldman L, Brann DW, Simone D, Mahesh VB (1997). Progesterone Metabolite allopregnanolone in women with premenstrual syndrome. Obstet Gynecol 90: 709–714.
Shope JT, Copeland LA, Dielman TE (1994). Measurement of alcohol use and misuse in a cohort of students followed from grade 6 through grade 12. Alcoholism 18: 726–733.
Torres JM, Ruiz E, Ortega E (2001). Effects of CRH and ACTH administration on plasma and brain neurosteroid levels. Neurochem Res 26: 555–558.
Acknowledgements
We thank Roussel–Uclaf for their generous gift of allopregnanolone reagent kits. We thank R Davies for revising the English text, and M Quintana and R Arcas for their technical assistance. The authors are indebted to the Clinico University and Trauma Hospitals of Granada for their help. This work was funded in part by DGICYT PM97-0177 and by the Andalusia Regional Government through its ‘Endocrinology and Metabolism’ research project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Torres, J., Ortega, E. Alcohol Intoxication Increases Allopregnanolone Levels in Female Adolescent Humans. Neuropsychopharmacol 28, 1207–1209 (2003). https://doi.org/10.1038/sj.npp.1300170
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1300170
Keywords
This article is cited by
-
Pregnenolone effects on provoked alcohol craving, anxiety, HPA axis, and autonomic arousal in individuals with alcohol use disorder
Psychopharmacology (2023)
-
Sex differences in stress-induced alcohol intake: a review of preclinical studies focused on amygdala and inflammatory pathways
Psychopharmacology (2022)
-
GABAA receptor polymorphisms in alcohol use disorder in the GWAS era
Psychopharmacology (2018)
-
Effect of nucleus accumbens shell infusions of ganaxolone or gaboxadol on ethanol consumption in mice
Psychopharmacology (2015)
-
Variation in AKR1C3, which encodes the neuroactive steroid synthetic enzyme 3α-HSD type 2 (17β-HSD type 5), moderates the subjective effects of alcohol
Psychopharmacology (2014)
